
运动通过改善PD模型小鼠纹状体D2MSN-D1MSN侧抑制效应调节基底神经节信息输出
2020-03-25 03:07刘晓莉乔德才
体育科学 2020年1期
赵 刚,刘晓莉,乔德才
(北京师范大学 体育与运动学院,北京100875)
帕金森病(Parkinson’s disease,PD)是一种由于黑质-纹状体环路多巴胺(dopamine,DA)能神经元凋亡导致的神经退行性疾病,基底神经节直接/间接通路失衡引起的机体运动功能障碍是PD 的主要病理特征。纹状体是皮层信息输入基底神经节的主要核团,也是基底神经节的中央加工区,约95%的神经元为中等多棘神经元(medium spiny neuron,MSN),根据纹状体细胞膜上DA 受体类型可分为表达D1R 调控直接通路的MSN(D1-MSN)和表达D2R 调控间接通路的MSN(D2-MSN)(Gagnon et al.,2017)。新近神经解剖学的研究发现,在纹状体核团内不同类型的MSNs 高度叠加,相邻的MSN 之间形成γ-氨基丁酸(γ-aminobutyric acid,GABA)能突触连接,被称为侧抑制突触,并对直接/间接通路产生侧抑制效应(Faust et al.,2016)。PD 患者静止性震颤等精细运动功能障碍均可能与纹状体MSN 侧抑制效应的异常有关(Bian et al.,2015)。
在2008年,Taverna 等(2008)证实PD 动物模型存在纹状体D2-MSN 侧抑制突触连接减弱的现象。PD 病理状态下这种树突棘的减少被认为是导致基底神经节功能异常的解剖学基础(Surmeier et al.,2014)。本实验室前期研究已经证实,4 周跑台运动可缓解PD 小鼠纹状体D2-MSN树突棘脱落的现象(陈平,2018);纹状体D2-MSN 树突棘的运动依赖性重塑不仅在改善PD 小鼠皮层-纹状体突触传递和D2-MSN 兴奋性方面具有一定调节作用,而且也可能是重塑PD 小鼠纹状体D2-MSN 侧抑制突触连接的结构基础(赵刚 等,2019a)。为此,本研究拟采用光刺激靶向性操控D2-MSN 兴奋性,探讨D2-MSN 对D1-MSN 的侧抑制效应对基底神经节信息输出的影响,进一步从突触及通路水平上解释D2-MSN 在运动改善PD 小鼠基底神经节信息输出中的作用机制。
1 材料与方法
1.1 实验动物与分组
4 周龄雄性D2-Cre 转基因小鼠,由北京师范大学脑与认知研究院提供。小鼠分笼饲养,饲养过程中自然光照,保持自由进食饮水,室温22℃±2℃,相对湿度40%~60%。所有动物实验方案均经北京师范大学动物伦理委员会批准,并按照实验动物使用的3R 原则,对实验动物给予人道主义关怀。小鼠适应性饲养1 周,随机分为对照组(D2-Cre)和模型组,模型组经鉴定符合PD 模型的小鼠再随机分为PD 组(PDD2-Cre)和PD 运动组(PD+ExD2-Cre),每组各12 只(6 只用于电生理学实验,6 只用于行为学和免疫组织化学实验)。
1.2 光敏感蛋白转染及鉴定
病毒注射前24 h 小鼠禁食,自由饮水。术前通过腹腔注射5%水合氯醛(350 μg/g)麻醉,使用立体定位仪定位,在小鼠纹状体右侧(AP:+0.5 mm;R:1.8 mm;DV:-3.0 mm)注入兴奋性光敏感蛋白病毒(rAAV-Ef1α-DIOChR2-EYFP-WPRE-pA,枢密,武汉)(滴度≥2.00E+12 vg/ml)1 μl,以0.1 μl/min 的速度注射完成后留针3 min,缓慢移出注射器,缝合创面,并于缝合处涂抹青霉素预防感染,待小鼠清醒后单笼饲养。视紫红质通道蛋白(Channelrhodopsin-2,ChR2)是一种受光脉冲控制的兴奋性阳离子通道蛋白,表达ChR2 蛋白的细胞在蓝光刺激下使通道打开,Na+、Ca2+等进入细胞,产生一内向电流,引起细胞去极化,从而兴奋神经元。
1.3 PD模型小鼠制备、评价及运动干预方案
光敏感蛋白转染2 周后,腹腔注射5%水合氯醛麻醉(350 μg/g),将小鼠固定在立体定位仪上,沿颅顶矢状缝切开皮肤,在右侧纹状体(AP:+0.5 mm,R:1.8 mm)钻孔,用微量注射器(Legato,KDS,美国)在颅骨下-3.0 mm 和-2.0 mm 不同深度分别注入6-羟基多巴胺(6-Hydroxydopamine hydrobromide,6-OHDA,Sigma,美国)溶液(2 μg/μl,含0.02%抗坏血酸)4 μl,以0.5 μl/min 的速度注射完成后留针3~5 min,匀速退出进样器,缝合处理创面。6-OHDA 为选择性儿茶酚胺类神经化学损毁剂,进入体内后易氧化生成羟自由基和醌类等神经毒物,特异性地损毁黑质致密部DA 能神经元,引起DA 生成减少,黑质-纹状体DA系统功能退变,一般用于制备稳定有效的PD 动物模型。
6-OHDA 注射后第7 天及4 周运动干预后24 h,各组小鼠腹腔注射APO(0.5 μg/g)诱导旋转行为,记录30 min内小鼠的旋转圈数。以向损伤侧旋转圈数与向健侧旋转圈数的差值作为小鼠净旋转圈数,净旋转圈数>120 r/30 min 作为PD 小鼠模型制备成功的标准,剔除未达到标准的小鼠(于燕 等,2015)。
运动干预方案采用匀速跑台运动(18 m/min,40 min/天,连续5 天/周,共4 周),运动强度约为70%~76%V·O2max(Høydal et al.,2007)。PD+ExD2-Cre组 在6-OHDA 注 射1 周后开始运动干预;非运动组小鼠置于跑台中相同时间,但不参与运动。
1.4 离体脑片制备
所有实验结束后,对各组小鼠腹腔注射水合氯醛麻醉(5%,500 μg/g),断头取全脑,在0℃氧饱和的改良人工脑脊液(modified artificial cerebrospinal fluid,mACSF)中修块。使用高速振动切片机(VT1200S,Leica,德国)对修块后的脑组织纹状体进行冠状切片(厚度400 μm,切片过程中持续通氧)。将脑片在人工脑脊液(artificial cerebro spinal fluid,ACSF)孵育槽中孵育1 h 以上备用,孵育过程中保持31℃并持续通入混合气体(95% O2/5% CO2)。
1.5 全细胞膜片钳记录
将脑片固定于记录槽中央(灌流氧饱和的ACSF),利用显微镜微分干涉相差成像系统和红外COD 摄像机(显微镜选择荧光模式),在视野中选取有荧光反应且状态良好的纹状体D2-MSN,以此为视野中心,反复切换荧光和普通模式寻找状态良好、不具有荧光反应的D1-MSN。操作机械臂将记录电极移入记录槽中,入液阻抗约为5~9 MΩ。给予记录电极正气压,移动电极将电极尖端压至D1-MSN 胞体表面,观察到胞体压痕并且电极阻抗上升后释放正压并持续给予低强度负压;同时给予-70 mV 钳制电压,电极阻抗逐渐上升高于1 GΩ 时形成高阻封接。记录波形趋于平缓,封接状态稳定后给予强而短促的负压或通过放大器给予Zap 电流刺激,破膜形成全细胞记录(马婧 等,2018)。
所有原始信号都通过膜片钳放大器(MultiClamp 700B,Molecular Devices,美国)检测、放大,1 KHz Bessel低通滤波,再经数模转换器(Digidata 1550A,Molecular Devices,美国),导入pClamp 10.5 软件记录。以串联电阻≤25 MΩ,静息膜电位≤-70 mV,且整个记录过程中变化幅度<20%为标准判断是否进行后续膜片钳实验(马婧,2015)。
动作电位的记录(action potential,AP):在电流钳模式下,给予D1-MSN 步阶变化的刺激电流,使其爆发动作电位。给予刺激电流(500 ms,0.1 Hz,-100~500 pA,以每20 pA 的步阶递增),统计随着注入电流的增大,D1-MSN爆发动作电位的放电频率。通过473 nm 蓝色激光刺激视野中心位置的D2-MSN,统计光刺激后D1-MSN 动作电位的发放频率。
光刺激诱发的抑制性突触后电流(optogenetic stimulation evoked inhibitory postsynaptic currents,oIPSC):ACSF加入CNQX(10 μM,谷氨酸受体AMPAR 拮抗剂)和AP5(50 μM,谷氨酸受体NMDAR 拮抗剂),将细胞膜电位钳制在-55 mV。待药物起作用后,稳定记录D1-MSN IPSC 5 min 作为基线数据。通过473 nm 蓝色激光照射视野中心位置的D2-MSN(50 ms on/50 ms off),稳定记录D1-MSN oIPSC 5 min(baseline,base)后向ACSF 中加入多巴胺II 型受体(dopamine type II receptor,D2R)激动剂喹吡罗(5 μM ,quinpirole,Quin),稳定记录oIPSC 5 min。通过循环泵加入ACSF(含CNQX 10 μM 和AP5 50 μM)快速洗脱,稳定记录oIPSC 5 min,统计加入Quin 后oIPSC 幅值与基线base 的比值。
1.6 纹状体-黑质网状部抑制性突触后场电位的记录
刺激电极置于纹状体背外侧,玻璃记录电极置于黑质网状部(substantia nigrapars reticular,SNr),ACSF 需加入CNQX(10 μM)以及AP5(50 μM),阻断Glu 介导的兴奋性电流,记录纹状体-黑质网状部通路的抑制性突触后场电位(field inhibitory post synaptic potential,fIPSP)。给予强度递增的电流刺激,获得纹状体产生fIPSP 的最小刺激强度,并增加刺激强度获得最大fIPSP 幅值;每30 s 给予1 个该强度的刺激,稳定记录15 min 基线;基线稳定后,刺激强度维持不变,同步给予纹状体脑区D2-MSN 473 nm 蓝光刺激(50 ms off/50 ms on),记录5 组fIPSP,取其平均值。
1.7 纹状体络氨酸羟化酶表达水平检测
小鼠腹腔注射5%水合氯醛麻醉(350 μg/g),经左心室-升主动脉插管灌流(30 ml 生理盐水和30 ml 4%多聚甲醛溶液),灌流结束后迅速取出脑组织置于4%的多聚甲醛溶液中固定24 h。将脑组织取出后置于30%的蔗糖溶液中进行脱水后修块、包埋,修整并进行冠状切片,每只6张脑片切片(10 μm)。对脑片切片进行免疫组织化学染色,按照1:1 000 稀释加入一抗(兔多克隆抗体,Abcam,美国),按照1:200 稀释加入二抗(过氧化物标记的抗兔IgG抗体,KPL,美国),荧光显微镜(DP72,Olympus,美国)对纹状体背侧区域拍照。采用Image-Pro Plus 6.0 软件对络氨酸羟化酶(tyrosine hydroxylase,TH)免疫阳性纤维光密度(optical density,OD)进行统计分析。
1.8 数据处理
所有电生理数据均以Mean±SEM 表示,其余行为学及免疫组织化学数据均以M±SD表示。利用pClamp 10.5软件分析oIPSC 数据,统计oIPSC 的幅值。利用SPSS 20.0软件采用单因素方差分析和卡方检验分析各指标的组间差异,P<0.05 表示有统计学意义。
2 研究结果
2.1 PD小鼠模型及纹状体光敏感蛋白转染情况鉴定
本实验中共用38 只小鼠采用纹状体同一位点、不同深度注射6-OHDA 的方法建立小鼠PD 模型,符合PD 模型标准的小鼠共24 只,成模率为63.15%。成模小鼠APO 旋转实验结果为189.23±16.46 r/30 min,净转数大于120 r/30 min。4 周运动干预后各组小鼠APO 诱导的旋转圈数结果如图1B 所示,D2-Cre 组小鼠净旋转圈数为35.16±8.76 r/30 min,PDD2-Cre组小鼠净旋转圈数为184.23±11.26 r/30 min,PD+ExD2-Cre组小鼠净旋转圈数为166.5±7.96 r/30 min;PDD2-Cre组小鼠净旋转圈数较D2-Cre 组显著增加(P<0.01),PD+ExD2-Cre组净旋转圈数较PD 组显著降低(P<0.05),但仍显著高于D2-Cre 组(P<0.01)。

图1 各组小鼠APO诱导的旋转圈数统计Figure 1.The Numbers of APO-induced Rotations in Each Group
免疫组织化学结果显示,PDD2-Cre及PD+ExD2-Cre组小鼠损毁侧(右侧)纹状体TH 阳性纤维出现严重丢失(图2A)。PDD2-Cre及PD+ExD2-Cre组小鼠纹状体损伤侧/未损伤侧TH 阳性纤维表达比值显著低于D2-Cre 组(P<0.05),PDD2-Cre及PD+ExD2-Cre组之间无显著性差异(P>0.05,图2B)。
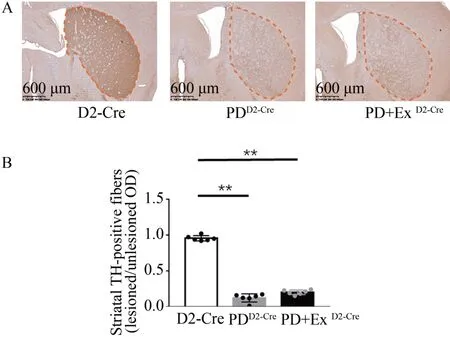
图2 各组小鼠纹状体TH阳性纤维表达Figure 2.Striatal TH Positive Fiber Expression in Each Group
对D2-Cre 小鼠转染兴奋性光敏感蛋白病毒(rAAVEf1α-DIO-ChR2-EYFP-WPRE-pA)共聚焦扫描结果显示,在绿色激发光下,D2-Cre 小鼠纹状体D2-MSN 出现明显绿色荧光(图3B);通过给予脑片蓝光刺激,在膜片钳显示器中可以观察到明显反应(图3C)。表明,将兴奋性光敏感蛋白病毒成功转染至纹状体D2-MSN。

图3 纹状体光敏感蛋白转染情况Figure 3.Optogenetic Viral Expression in Str
2.2 光刺激PD 模型小鼠纹状体D2-MSN 对D1-MSN 动作电位发放的影响
采用全细胞膜片钳结合光遗传技术,在荧光显微镜下特异性区分D2-MSN 和D1-MSN(图4A、B),钳制D1-MSN并注入一系列电流,使其爆发动作电位,D1-MSN 表现出内向整流和动作电位发放延迟等MSN 典型的电生理特性(图4C、D、E);用473 nm 的蓝色激光刺激视野中心D2-MSN,观察激活D2-MSN 对D1-MSN 动作电位发放的影响。统计结果显示,注入300 pA 电流时,D1-MSN 动作电位发放个数:D2-Cre 组 为15.43±0.88 个,PDD2-Cre组 为16.33±1.01 个,PD+ExD2-Cre组为15.97±1.08 个,各组间无显著性差异(P>0.05)。注入300 pA 电流并同时给予473 nm 的蓝色激光刺激视野中心D2-MSN 时,D1-MSN 动作电位发放个数出现明显变化,D2-Cre 组为7.1±0.66 个,PDD2-Cre组为4.11±0.55 个,显著低于D2-Cre 组(P<0.05);PD+ExD2-Cre组为5.75±0.4 个,显著高于PD 组,但低于D2-Cre 组(P<0.05)(图4F)。
2.3 D2R 激动剂对PD 模型小鼠D2MSN-D1MSN 抑制性效应的影响
D2R 激动剂干预对光刺激D2-MSN 诱导的D1-MSN oIPSC 的影响,各组小鼠存在一定差异(图5)。统计结果显示,D2-Cre 组小鼠纹状体D1-MSN oIPSC 的幅值与基线的比值为56.25%±2.98%(n=8 cells),PDD2-Cre组小鼠为79.87%±2.68%(n=8 cells),PD+ExD2-Cre组小鼠为71.12%±5.38%(n=8 cells);PDD2-Cre组小鼠oIPSC 幅值与基线的比值显著高于D2-Cre 组(P<0.05),PD+ExD2-Cre组小鼠显著低于PDD2-Cre组,但高于D2-Cre 组(P<0.05,图5D)。
2.4 光刺激D2-MSN 对PD 模型小鼠纹状体-黑质网状部抑制性突触后场电位的影响
采用离体脑片场电位结合光遗传技术,观察光刺激D2-MSN 诱导各组小鼠SNr 的fIPSP 最大幅值变化情况。结果显示,D2-Cre 组fIPSP 最大幅值为1.38±0.12 mV(n=7 slices),PDD2-Cre组fIPSP 最大幅值为1.02±0.17 mV(n=6 slices),PD+ExD2-Cre组fIPSP 最大幅值为1.18±0.07 mV(n=7 slices);PDD2-Cre组小鼠fIPSP 最大幅值显著低于D2-Cre 组(P<0.05),PD+ExD2-Cre组小鼠fIPSP 最大幅值显著高于PDD2-Cre组,但低于D2-Cre 组(P<0.05)(图6C、D)。
3 讨论与分析
纹状体是基底神经节中运动调控的重要脑区,激活纹状体D1-MSN,调控直接通路,可易化运动行为;激活纹状体D2-MSN,调控间接通路,则抑制运动行为。由于D1MSN 和D2MSN 在纹状体内的分布高度重叠,很难依靠其解剖形态或电生理特性进行有效区分。随着光遗传学技术的问世,对D1-MSN 和D2-MSN 神经元特异性识别及操控成为可能(Nagel et al.,2002;Zhang et al.,2007)。利用光遗传学、病毒转染等技术的研究进一步发现,纹状体相邻MSN 之间存在旁支突触结构,称之为侧抑制突触;并根据突触前后胞体表达的DA 受体类型分为D2MSND2MSN、D2MSN-D1MSN、D1MSN-D2MSN 及 D1MSND1MSN 4 种突触连接方式。侧抑制突触不仅对突触后神经元产生侧抑制效应,还可对突触前神经元产生前馈抑制,进而调节直接或间接通路的活性。
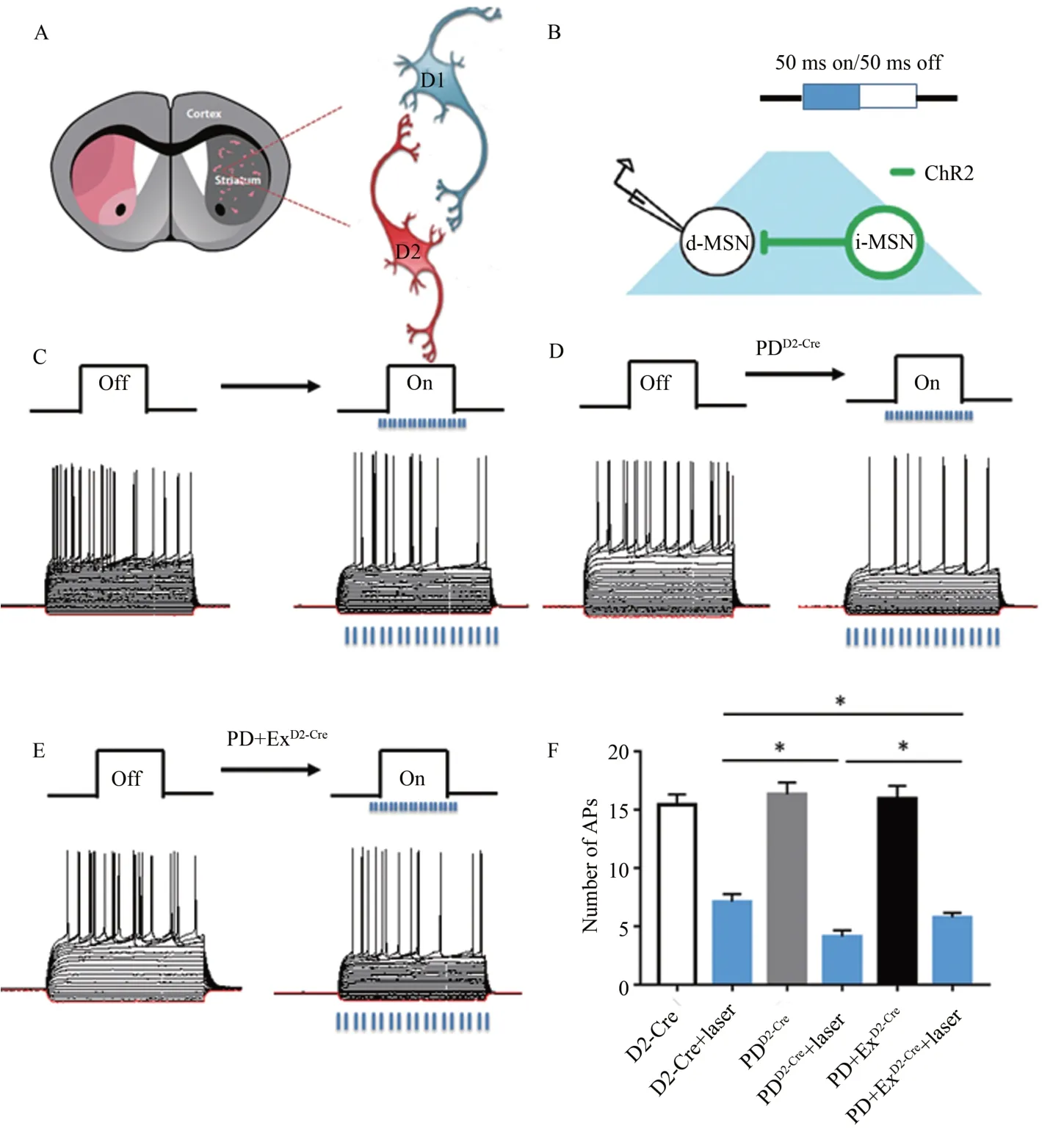
图4 光刺激前后各组小鼠D1-MSN的动作电位发放情况Figure 4.The APs of D1-MSN before and after Optogenetic Stimulation in Each Group
3.1 运动降低PD模型小鼠D2MSN-D1MSN侧抑制效应
光遗传结合全细胞膜片钳技术已运用于PD 病理机制的研究中。Cui 等(2013)利用光遗传学技术兴奋小鼠纹状体的D2-MSN,成功使小鼠表现出PD 类似的行为功能障碍。Dobbs 等(2016)采用光遗传技术激活表达有ChR2的小鼠D2-MSN,抑制了D1-MSN 兴奋性,初步证实在纹状体MSN 之间存在侧抑制效应,并可能参与直接/间接通路信号调控的假说。Taverna 等(2008)观察到PD 小鼠模型存在MSN 侧抑制突触连接减弱的现象,且这种现象主要发生在D2-MSN。Wei 等(2017)进一步使用膜片钳配对记录的方法研究发现,PD 病理状态下D2-MSN 侧抑制突触的IPSC 幅值升高,侧抑制效应发生异常。本实验中对D2-Cre 小鼠转染兴奋性光敏感蛋白病毒(rAAV-Ef1α-DIOChR2-EYFP-WPRE-pA),通过473 nm 蓝光刺激D2-MSN,发现各组小鼠相邻D1-MSN 动作电位发放频率和数量均显著降低,证实D2-MSN 与相邻D1-MSN 存在侧抑制效应;且PD 组小鼠侧抑制突触的oIPSC 幅值显著升高,侧抑制效应发生异常,与Wei 等(2017)研究结果一致。本实验中,PD+ExD2-Cre组小鼠在给予D2-MSN 473 nm 蓝光刺激后相邻D1-MSN 动作电位发放数量显著高于PD 组,但低于D2-Cre 组,表明运动干预减弱了PD 小鼠纹状体D2MSND1MSN 的侧抑制效应,其机制可能与运动改善了PD 小鼠皮层-纹状体突触传递异常,降低了D2-MSN 的兴奋性,减少了侧抑制突触前GABA 的释放有关(赵刚 等,2019b)。本实验室前期研究已经证实,运动干预可选择性逆转PD动物D2-MSN 的树突棘丢失,使纹状体突触结构发生运动依赖性重塑(陈平,2018),这一结果也为运动改善D2MSN-D1MSN 侧抑制效应提供了神经解剖学基础。

图5 D2R激动剂干预对纹状体D2MSN-D1MSN oIPSC的影响Figure 5.The Effect of D2R Agonist on D2MSN-D1MSN oIPSC
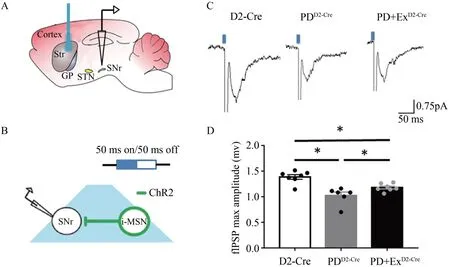
图6 光刺激D2-MSN对SNr的fIPSP最大幅值的影响Figure 6.The Effect of Optogenetic Stimulation D2-MSN on SNr fIPSP
3.2 D2R 在运动改善PD 模型小鼠D2MSN-D1MSN 侧抑制效应中的作用
纹状体D2MSN-D1MSN 侧抑制效应的强弱可由突触前与突触后两条途径调节。DA 受体主要参与侧抑制突触前神经元兴奋性的调节。当突触前MSN 细胞膜上D1R激活时,胞体内环磷酸腺苷(Cyclic Adenosine monophosphate,cAMP)水平上调,进而增加蛋白激酶A 水平,从而增强G 蛋白耦联受体活性,使更多GABA 进入囊泡释放至突触间隙;相反,当突触前MSN 细胞膜上D2R 激活时,胞体内cAMP 水平下调,减少GABA 的释放。Lalchandani等(2013)利用光遗传技术将ChR2 转染在小鼠D2-MSN,发现给予D2R 激动剂Quin 干预可减弱光刺激D2-MSN 对D1-MSN 的IPSCs,但不影响间接通路中接受D2-MSN 投射的苍白球外侧部(external segment of globus pallidus,GPe)神经元的IPSCs。本实验中,通过给予D2R 激动剂Quin 抑制突触前D2-MSN 胞体的兴奋性,显著降低D2MSN-D1MSN 侧抑制效应。提示,D2R 通过降低D2-MSN 兴奋 性 减 少GABA 释放,调节D2MSN-D1MSN 侧抑制效应。
目前,有关运动干预对PD 小鼠纹状体侧抑制效应影响的研究鲜见相关报道。本实验采用光遗传、药理学结合离体脑片膜片钳技术研究发现,给予D2R 激动剂Quin干预后,光刺激D2-MSN 诱导的D1-MSN oIPSC,PD+ExD2-Cre组小鼠显著低于PD 组,表明D2R 在运动改善纹状体D2MSN-D1MSN 侧抑制效应的调节中具有重要作用。本实验室前期的研究证实,运动的神经保护作用可上调纹状体D2R 表达水平(王弘,2016),抑制PD 动物D2-MSN的兴奋性。由此推测,运动干预减弱PD 小鼠纹状体D2MSN-D1MSN 侧抑制效应异常与侧抑制突触前膜D2R功能改变有关,D2R 可能是D2MSN-D1MSN 侧抑制效应调节的重要细胞分子。
3.3 运动通过改善PD 模型小鼠D2MSN-D1MSN 侧抑制效应调节SNr 信号输出
SNr 是基底神经节直接与间接通路最终信息整合与输出的核团之一,可接收多种兴奋或抑制性神经投射。神经解剖学研究证实,纹状体MSN 之间不仅存在短侧支突触连接,还有长距离轴突投射至SNr。Schmidt 等(2013)研究发现,无论是否成功停止动作,丘脑底核(subthalamicnucleus,STN)神经元都出现较高的兴奋性,但只有当真正成功停止动作时,SNr 神经元才会出现较高的兴奋性,表明快速反应动作终止需要侧抑制效应参与。此外,Gulley 等(2002)发现,大鼠的SNr 会在抑制性行为的开始阶段出现放电频率降低,而此时直接通路其他核团的神经元仍处于兴奋状态。提示,来自纹状体D2MSND1MSN 的侧抑制效应可能也参与SNr 信息输出的调节。
本研究中向ACSF 中加入CNQX(10 μM)及AP5(50 μM),阻断Glu 介导的兴奋性电流,在SNr 记录到的fIPSP主要为纹状体D1-MSN GABA 能神经元介导。研究结果表明,光刺激纹状体D2-MSNPD,SNr 的fIPSP 最大幅值显著降低,与前一实验PD 组小鼠D1-MSN oIPSC 幅值升高相对应。提示,PD 病理状态下D2MSN-D1MSN 侧抑制效应增强对SNr 信息输出产生抑制作用。4 周运动干预增强了PD 小鼠SNr 的fIPSP 最大幅值,可能与运动降低D2MSN-D1MSN 侧抑制效应,提高D1-MSN 的兴奋性,加强了SNr 的信息输出有关。
侧抑制效应对基底神经节输出核团的调控机制仍存在争议。有学者认为,来自皮层的信息经过纹状体MSN处理后,大部分沿直接与间接通路投射至苍白球内侧部/黑质网状部复合体(GPi/SNr),此时相邻MSN 通过侧抑制效应对部分信息进行调节,并沿轴突投射传至GPi/SNr;生理状态下,GPi/SNr 通过以上两种方式接收的兴奋/抑制信息输出具有较高的对比度,可将清晰的信息反馈回皮层精确调控运动(Wei et al.,2017,图7)。本研究认为,PD病理状态下,纹状体DA 投射减少,D2R 表达下调及出现超敏,MSN 侧抑制突触前膜GABA 释放减少,侧抑制效应异常,此时GPi/SNr 接收的兴奋与抑制信息输出的对比度降低,无法向皮层精确反馈动作信息,引起运动功能障碍。运动可能通过上调D2R 表达,改善D2MSN-D1MSN侧抑制效应,加强GPi/SNr 兴奋与抑制信息输出的对比度,提高向皮层反馈动作信息的精确性,改善PD 运动功能障碍。
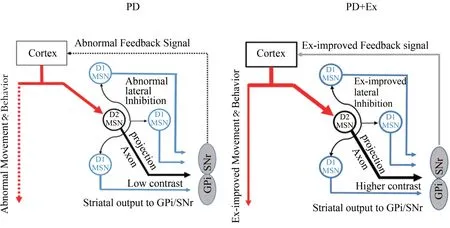
图7 D2MSN-D1MSN侧抑制效应对SNr信号输出的调节Figure 7.The Adjustment Diagram of D2MSN-D1MSN Lateral Inhibition on SNr Signal Output
4 结论
PD 小鼠纹状体D2MSN-D1MSN 侧抑制效应异常,并影响了SNr 的信息输出,这可能是导致PD 基底神经节功能紊乱的重要原因之一。4 周运动干预通过改善PD 小鼠纹状体D2MSN-D1MSN 侧抑制效应,调节了SNr 的信息输出。D2R 在运动改善PD 小鼠纹状体D2MSN-D1MSN 侧抑制效应调节基底神经节信息输出中起重要作用。
猜你喜欢
介入放射学杂志(2022年8期)2022-11-26
中草药(2022年17期)2022-09-05
云南中医中药杂志(2022年5期)2022-05-18
健康之家(2021年19期)2021-05-23
电子产品世界(2021年8期)2021-01-16
阅读(科学探秘)(2019年5期)2019-07-19
教育教学论坛(2019年18期)2019-06-17
中国计算机报(2019年49期)2019-02-07
中西医结合心血管病电子杂志(2018年28期)2018-11-19
中国新闻周刊(2017年36期)2017-10-21
