
田间禾本科杂草对水稻条纹病毒流行风险评估调查
2020-03-24 11:22王艺晓朴君程兆榜邓金花周益军李硕
杂草学报 2020年4期
关键词:风险评估
王艺晓 朴君 程兆榜 邓金花 周益军 李硕

摘要:采用室内灰飞虱接种鉴定的方法评价了江苏地区主要杂草对水稻条纹病毒(rice stripe virus, 简称RSV)的感染率。结果显示,看麦娘、稗、旱稗、狗尾草、升马唐的感染率较高,为38.24%~62.00%,具有成为RSV“桥梁”寄主的可能性。对病害大流行年份(2005年)江苏主要杂草种类RSV感染率田间调查工作进行总结,发现野生禾本科杂草中存在RSV感染,但其感染率低于同期小麦植株的感染率,这说明小麦是RSV侵染循环中重要的“桥梁”寄主。于病害不流行年份(2018—2019年)在江苏多地采集田边稗草和狗尾草,经分子生物学检测,可检测出RSV。禾本科杂草对病害流行的贡献度低于小麦,其在病害侵染循环中的作用主要体现在维持RSV在农田生态系统中的存在。
关键词:禾本科杂草;水稻条纹病毒;病毒感染率;田间调查;风险评估
中图分类号:S451文献标志码:A文章编号:1003-935X(2020)04-0014-06
Risk Assessment of Gramineae Weeds for Rice Stripe Virus Epidemic in the Field
WANG Yi-xiao1,2, PIAO Jun1, CHENG Zhao-bang2, DENG Jin-hua2, ZHOU Yi-jun2, LI Shuo2
(1.School of Life Science,Liaoning Normal University, Dalian 116081, China;2.Institute of Plant Protection,Jiangsu Academy of Agricultural Sciences,Nanjing 210014,China)
Abstract:The infection rate of rice stripe virus (RSV) to main weeds in Jiangsu Province was evaluated using Laodelphax striatellus for inoculation in laboratory. Alopecurus aequalis, Echinochloa crusgalli, E. hispidula, Setaria viridis and Digitaria ciliaris could serve as “bridge” hosts of RSV. Their infection rates were between 38% and 62%. RSV infection rate to main weed species in fields in Jiangsu Province during the epidemic year (2005) was summarized. RSV infection existed in wild Gramineae weeds, but its infection rate was lower than that in wheat plants at the same time. It suggested that wheat was an important “bridge” host in RSV infection cycle. RSV was detected in E. crusgalli and S. viridis collected from many locations in Jiangsu Province when the disease was not prevalent (2018—2019). The contribution of Gramineae weeds to the disease epidemic was lower than that of wheat, and their role in the disease infection cycle was mainly reflected in maintaining the existence of RSV in the farmland ecosystem.
Key words:Gramineae weed; rice stripe virus; virus infection rate; field survey; risk assessment
水稻條纹叶枯病是水稻重要病毒病害之一,该病由介体昆虫灰飞虱(Laodelphax striatellus Fallén)传播水稻条纹病毒(rice stripe virus, 简称RSV)引起。在2003—2010年,水稻条纹叶枯病在以江苏省为中心的华东稻区暴发,对水稻生产造成了极大损失[1-2]。病害曾在我国扩及到18个省(市、自治区)的多个稻区,其中以江苏、浙江、安徽、河南、山东、云南等地的粳稻田发病最为普遍[1]。此外,该病害在东亚地区的日本、韩国、朝鲜等地也有分布[3-4]。灰飞虱是RSV的主要传播介体,它与水稻条纹叶枯病的流行和暴发有着非常密切的关系[1,5]。RSV可由灰飞虱经卵传至子代[6-7],这一特性是RSV在稻麦轮作区多年持续流行的重要原因,所以条纹叶枯病一旦大发生,就可以造成持续数年的流行危害。2010年以后,随着以抗病品种和“防虫网阻断传毒”为核心技术的绿色综合防控体系大面积推广,水稻条纹叶枯病在华东稻区的感染趋势逐年下降[2,8]。近年来,该病主要在云南省流行危害,已成为当前危害云南水稻最严重的病毒病[9],并向四川省南部地区扩散[10]。
RSV为纤细病毒属(Tenuivirus)成员,自然条件下只侵染禾本科植物,与同属的其他病毒相比,RSV的寄主范围相对较广[11]。实验室条件下,RSV可以通过摩擦接种侵染本氏烟[12],也可通过灰飞虱传毒侵染拟南芥[13]。研究人员室内接种鉴定发现,除水稻外,RSV还能侵染大麦、小麦、看麦娘(Alopecurus aequalis Sobol.)、玉米、早熟禾等80多种禾本科作物和杂草[4,14],故RSV在田间存在着广泛的潜在寄主。在江苏省稻麦轮作区,大量种植的冬小麦成为病毒和灰飞虱的主要越冬场所和中间寄主[15]。虽然RSV的禾本科杂草寄主范围已有详细报道,但各杂草种类对RSV的感染率,尤其是田间自然条件下的感染率还不清楚。条纹叶枯病在江苏大流行期间,马唐[Digitaria sanguinalis (L.) Scop.]、稗[Echinochloa crusgalli (L.) Beauv.]、升马唐[Digitaria ciliaris (Retz.) Koel.]等禾本科杂草在田间已普遍发生,这些杂草有作为RSV流行过程中“桥梁”寄主的潜力,为介体传毒提供毒源,但它们在病害发生流行中的作用尚不明确。评估调查杂草寄主在RSV流行中的风险作用,对解析RSV的侵染循环规律有着重要意义,同时也可以为水稻条纹叶枯病的综合防治提供参考。因此,本研究采用室内接种鉴定方法评价了江苏地区主要杂草种类对RSV的感染率,并对病害大流行年份(2005年)江苏主要杂草种类RSV田间感染率调查工作进行了总结,同时,于病害不流行年份(2018—2019年)在江苏多地采集田边杂草,进行RSV感染情况检测,综合分析杂草在病毒流行中的风险作用。
1 材料与方法
1.1 传毒介体
带毒(RSV)灰飞虱群体为笔者所在实验室筛选保存。灰飞虱采集自江苏省海安市,由江苏省农业科学院植保所植物病毒研究室利用RSV经卵传播特性经多代筛选获得高亲和性带毒(RSV)群体,并在实验室内常年维持带毒,具体筛选方法参照刘海建等的研究[16]。本试验所用灰飞虱群体带毒率为80%。
1.2 杂草寄主室内接毒及检测
于田间收集江苏地区主要禾本科杂草的种子,杂草种类包括看麦娘、牛筋草[Eleusine indica (L.) Gaertn.]、馬唐、狗尾草[Setaria viridis (L.) Beauv.]、稗[Echinochloa crusgalli (L.) Beauv.]、旱稗[Echinochloa hispidula (Retz.) Nees.]、升马唐、千金子[Leptochloa chinensis (L.) Nees.]和菵草[Beckmannia syzigachne (Steud.) Fern.],共计9种。杂草种子用赤霉素浸种 8 h,在烧杯中播种,置于26 ℃光照培养箱种。在苗期按照3头/株的比例接入带毒灰飞虱,接毒3 d后,杂草苗移至室外无虫网室生长,并观察记录叶片症状。每个品种设3个重复,以不接毒样本作为对照,2周后利用RSV单克隆抗体采用斑点酶联免疫吸附试验(Dot-ELISA) [17]检测杂草样本的感染率。
1.3 病害大流行年份江苏地区田间杂草RSV感染率调查
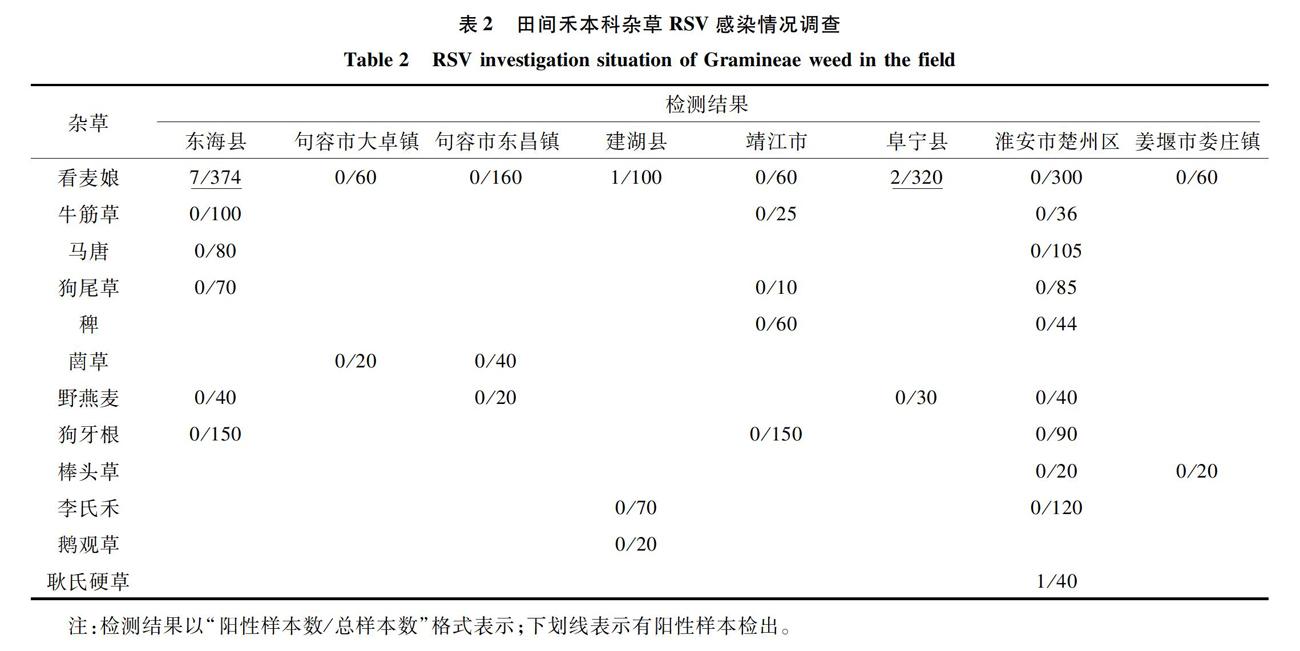
于RSV大流行年份(2005年)春季在江苏地区已发生RSV感染小麦的麦田中采集野生禾本科杂草。采集地点包括江苏省东海市、句容市大卓镇和东昌镇、建湖县、靖江市、阜宁县、淮安市楚州区、泰州市姜堰区娄庄镇。采集杂草种类为江苏地区麦田4—5月常见野生禾本科杂草,包括看麦娘、牛筋草、马唐、狗尾草、稗、菵草、野燕麦(Avena fatua)、狗牙根[Cynodon dactylon (L.) Pers.]、棒头草(Polypogon fugax Nees ex Steud.)、李氏禾[Leersia hexandra Swartz]、鹅观草(Roegneria kamoji Ohwi)和耿氏硬草[Sclerochloa kengiana (Ohwi) Tzvel.],共计12种。每种杂草样本数为10~374株不等,实验室内采用Dot-ELISA法检测RSV感染率,并对同期田间小麦病株率进行调查。
1.4 病害不流行年份江苏地区田间杂草RSV感染检测
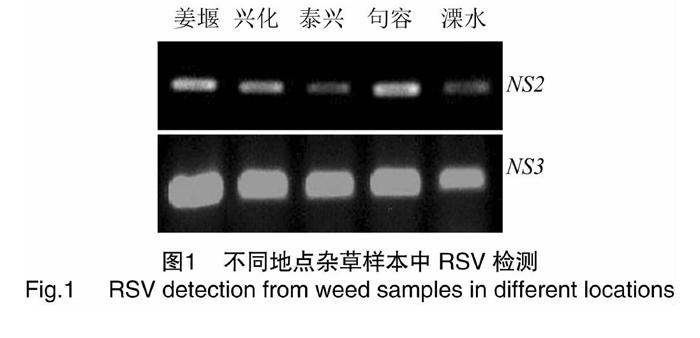
于病害不流行年份(2018—2019年)在江苏省泰州市姜堰区、兴化市、泰兴市、句容市以及南京市溧水区等地稻田田埂采集表现类似“花叶条纹”症状的稗草和狗尾草。用Trizol法提取杂草叶片总RNA,设计靶向RSV NS2和NS3基因的2对特异检测引物,ns2-F:5′-ATGGCATTACTCCTTTTCAAT-3′和ns2-R:5′-TCACATTAGAATAGGACACT-3′,ns3-F:5′-ATGAACGTGTTCACATCG-3′和ns3-R:5′-CCATGAAGAAGCCTCAGAAACTG-3′,进行逆转录-聚合酶链反应(RT-PCR)扩增,扩增产物经1%琼脂糖凝胶电泳分析,并回收测序。测序由生工生物工程(上海)股份有限公司完成。
2 结果与分析
2.1 杂草寄主室内接毒RSV检测结果
室内用带毒灰飞虱接种9种常见杂草,经 Dot-ELISA 检测,结果显示除菵草外,看麦娘、牛筋草、马唐、狗尾草、稗、旱稗、升马唐和千金子8种杂草均可被RSV侵染(表1),其中,看麦娘、升马唐、狗尾草、稗、旱稗感染率较高,达38.24%~62.00%,表明这几种杂草对RSV较为敏感,具有在RSV流行过程中作为“桥梁”寄主的可能性。
2.2 病害大流行年份田间杂草RSV感染率调查结果
于RSV大流行年份(2005年)在各地发生RSV感染的麦田中采集野生禾本科杂草样本,共计12种,2 865份。经Dot-ELISA检测,发现野生禾本科杂草中存在RSV感染,但感染率较低。采自东海市、建湖县和阜宁县的看麦娘中检测出RSV,感染率分别为1.87%、1.00%和0.63%,采自淮安市楚州区的耿氏硬草RSV检出率为2.50%,其余样本均未检出RSV(表2)。而同期田间小麦病株率调查结果(阜宁县)显示,发病田块病株率为3.33%~16.40%,平均病株率为6.95%。这些结果说明RSV在自然条件下可由介体灰飞虱传播侵染禾本科杂草,但杂草的感染率远低于小麦的感染率,小麦是RSV侵染循环中重要的“桥梁”寄主。
2.3 病害不流行年份田间杂草RSV感染检测结果
于病害不流行年份(2018—2019年)在江苏多地稻田边采集显示出类似“花叶条纹”症状的稗草和狗尾草样本。经RT-PCR检测,2对引物均可扩增出符合目的基因(NS2和NS3)预期大小的特异条带(图1),经测序表明所扩增基因与目的序列一致,说明于姜堰、兴化、泰兴采集的稗草和句容、溧水采集的狗尾草样本均出现RSV感染。由于样本采集田块附近的水稻植株未发生感病情况,这表明在病害不流行时期,禾本科杂草在病害侵染循环中的作用主要体现在维系RSV在农田生态系统中的存在。
3 讨论
2003—2010年,水稻条纹叶枯病在江苏地区暴发流行,给水稻生产造成严重威胁。在研究病害暴发流行规律时,笔者所在研究团队发现稻麦轮作栽培方式是导致毒源积累和介体灰飞虱大发生的重要原因[15]。麦田是RSV和灰飞虱的主要越冬场所,小麦是RSV病害循环的重要中间寄主[18],而田间禾本科杂草在病害侵染循环中的作用有待明确。本试验采用室内接种方法评价了江苏地区主要禾本科杂草对RSV的感染率,并对病害大流行年份江苏地区主要杂草种类的RSV田间感染率的调查工作进行了总结,同时对病害不流行年份田边杂草的病毒感染情况进行了检测。
室内灰飞虱接毒试验表明,看麦娘、升马唐、狗尾草、稗、旱稗对RSV感染率较高,具有成为RSV潜在“桥梁”寄主的可能性。但在春季麦田中调查禾本科杂草的病毒感染率发现,仅有少量植株为RSV阳性,低于小麦的病株率,这说明小麦可能是RSV侵染循环中重要的“桥梁”寄主。甚至在特定条件(如早播或春后出苗后毒源得到补充,灰飞虱群体带毒率提高)下,还会出现RSV严重危害小麦生长的现象[18-19],进一步促进病害大流行。禾本科杂草对病害流行的贡献度低于小麦,处于次要地位,可能原因如下:小麦作为麦田中的农作物被集中种植,相对于野生禾本科杂草为优势种群,加之农田除草措施进一步降低了杂草的数量;灰飞虱对小麦这类禾本科作物和禾本科杂草存在取食偏好性,小麦更满足灰飞虱的营养要求,同等条件下灰飞虱倾向于选择小麦作为寄主,只有小麦收割后灰飞虱才会被迫大量迁移到田边的野生杂草上取食;小麦可以为灰飞虱提供稳定的越冬场所,而很多禾本科杂草秋冬枯萎春季萌发,在早春无法及时为越冬带灰飞虱提供营养。上述原因综合作用导致灰飞虱传播RSV侵染禾本科杂草的概率较小。此外,室内接种鉴定发现,大多数禾本科杂草被灰飞虱传毒后,一般不容易出现花叶、褪绿等典型症状,这说明野生禾本科杂草对RSV抗性较高,这可能也是禾本科杂草田间感染率低的原因之一。
杂草在病害侵染循环中的作用主要体现在维系RSV在自然界中的存在。尽管RSV可以由灰飞虱经卵传播,但由于RSV对卵孵化的负效应[8],带毒群体存在自然衰减,在抗病品种大面积种植的条件下,灰飞虱从水稻植株重新獲毒的概率大大降低,近年来江苏多地田间灰飞虱带毒率为0就是1个例证。但RSV并没有消失,主要原因是杂草作为野生寄主,为RSV在农田系统中存活提供了条件。本试验于病害不流行的2018—2019年在多地稻田边采集显示“花叶条纹”症状的稗草和狗尾草样本,经分子检测,确认为RSV感染,为上述观点提供了直接证据。本试验显示,部分禾本科杂草在室内条件下对RSV有较高感染率,说明这些杂草具备成为RSV“桥梁”寄主的潜力。江苏地区稻田田埂及下茬麦田杂草群落结构调查显示,禾本科杂草占有绝对优势,其中就包括对RSV有较高感染率的看麦娘、稗、马唐等[20-22]。野生禾本科杂草为RSV和灰飞虱的越冬和越夏2个重要环节提供了便利。在水稻条纹叶枯病的综合防控中,麦田翻耕破坏介体越冬场所的技术被应用,野生杂草可为其提供越冬场所。在水稻秧田期,防虫网覆盖隔离措施被广泛应用[2-24],而此时小麦已完成收割,无法迁移到水稻秧田的灰飞虱将被迫迁移到田边沟坎的杂草中生存。这种情况下,禾本科杂草则成为病害侵染循环中的重要一环,对RSV的流行存在非常重要,在防控不到位的情况下,可能还会引起条纹叶枯病的发生。因此,在条纹叶枯病的防控工作中,不应忽视对杂草的管理。
参考文献:
[1]周益军. 水稻条纹叶枯病[M]. 南京:江苏科学技术出版社,2010.
[2]周益军,李 硕,程兆榜,等. 中国水稻条纹叶枯病的研究进展[J]. 江苏农业学报,2012,28(5):1007-1015.
[3]Otuka A. Migration of rice planthoppers and their vectored re-emerging and novel rice viruses in East Asia [J]. Frontiers in Microbiology,2013,4:309.
[4]Toriyama S. Rice stripe virus [J]. CMI/ABB Description of Plant Viruses,1983,269:15.
[5]Hibino H. Biology and epidemiology of rice viruses [J]. Annual Review of Phytopathology,1996,34:249-274.
[6]Huo Y,Liu W W,Zhang F J,et al. Transovarial transmission of a plant virus is mediated by vitellogenin of its insect vector [J]. PLoS Pathogens,2014,10:e1003949.
[7]Deng J H,Li S,Hong J,et al. Investigation on subcellular localization of rice stripe virus in its vector small brown planthopper by electron microscopy [J]. Virology Journal,2013,10:310.
[8]Li S,Wang S J,Wang X,et al. Rice stripe virus affects the viability of its vector offspring by changing developmental gene expression in embryos [J]. Scientific Reports,2015,5:7883.
[9]张水英,谭冠林,任国敏,等. 2013—2014年云南主要稻区病毒病调查及RSV的分子变异[J]. 中国水稻科学,2015,29(5):535-545.
[10]郑桂贤. RSV四川分离物p3基因原核表达及其影响本氏烟内源基因表达研究[D]. 重庆:西南大学,2018.
[11]Falk B W,Tsai J H. Biology and molecular biology of viruses in the genus tenuivirus [J]. Annual Review of Phytopathology,1997,36:139-163.
[12]Yao M,Zhang T Q,Zhou T,et al. Repetitive prime-and-realign mechanism converts short capped RNA leaders into longer ones that may be more suitable for elongation during rice stripe virus transcription initiation [J]. Journal of General Virology,2012,93:194-202.
[13]Sun F,Yuan X,Zhou T,et al. Arabidopsis is susceptible to rice stripe virus infections [J]. J Phytopathol,2011,159(11):767-772.
[14]阮义理,金登迪,许如银. 水稻条纹叶枯病毒的寄主植物[J]. 植物保护,1984,10(3):22-23.
[15]程兆榜,何敦春,陈全站,等. 单季稻小麦轮作区灰飞虱发生规律[J]. 应用昆虫学报,2013,50(3):706-717.
[16]刘海建,程兆榜,王 跃,等. 灰飞虱传递水稻条纹病毒研究初报[J]. 江苏农业学报,2007,23(5):492-494.
[17]王贵珍,周益军,陈正贤,等. 水稻条纹病毒单克隆抗体的制备及检测应用[J]. 植物病理学报,2004,34(4):302-306.
[18]程兆榜,邓金花,任春梅,等. 江苏小麦条纹病毒病的发生及其病原的RT-PCR分析[J]. 麦类作物学报,2007,27(6):1138-1142.
[19]Xiong R Y,Cheng Z B,Wu J X,et al. First report of an outbreak of rice stripe virus on wheat in China [J]. Plant Pathol,2008,57:397.
[20]李淑顺,张连举,强 胜. 江苏中部轻型栽培稻田杂草群落特征及草害综合评价[J]. 中国水稻科学,2009,23(2):207-214.
[21]左然玲. 稻麦轮作田杂草迁移动态规律研究及其调控技术[D]. 南京:南京农业大学,2007.
[22]王开金,强 胜. 江苏麦田杂草群落的数量分析[J]. 草业学报,2007,16(1):118-126.
[23]李粉华,孙国俊,季 敏,等. 灰飞虱传水稻病毒病综合防控技术应用[J]. 江苏农业科学,2015,43(2):137-139.
[24]徐小兰,丁桂珍,胡莲凤,等. 水稻条纹叶枯病与黑条矮缩病综合防控技术研究[J]. 现代农业科技,2012 (5):195-196.
收稿日期:2020-09-13
基金项目:江苏省农业科技自主创新资金 [编号:CX(20)3135]。
作者简介:王艺晓(1996—),女,辽宁铁岭人,硕士研究生,主要从事植物病毒与昆虫介体互作相关研究。E-mail:wyx19960105@126.com。
通信作者:李 碩,博士,副研究员,主要从事昆虫介体传播植物病毒机理及控制技术相关研究,E-mail:lishuo@jaas.ac.cn;周益军,博士,研究员,主要从事植物病毒病害相关研究,E-mail:yjzhou@jaas.ac.cn。
猜你喜欢
现代营销·学苑版(2016年10期)2016-12-12
合作经济与科技(2016年24期)2016-12-07
时代金融(2016年27期)2016-11-25
人间(2016年24期)2016-11-23
价值工程(2016年29期)2016-11-14
企业技术开发·中旬刊(2016年10期)2016-11-12
时代金融(2016年23期)2016-10-31
大众理财顾问(2016年8期)2016-09-28
企业导报(2016年10期)2016-06-04
