
水盐胁迫对玉米茎木质部水力特性的影响
2020-03-20 06:32:30徐建新王潜高阳孙景生侯泽林
灌溉排水学报 2020年1期
徐建新,王潜,高阳,孙景生,侯泽林
水盐胁迫对玉米茎木质部水力特性的影响
徐建新1,王潜1,高阳2*,孙景生2,侯泽林1
(1.华北水利水电大学,郑州 450000;2.中国农业科学院 农田灌溉研究所,河南 新乡 453002)
【】探讨水盐胁迫下玉米茎木质部水力特征、光合作用和生长指标的响应,深入探究水盐胁迫诱导茎木质部栓塞形成的机理。通过室内玉米桶栽试验,设置水分胁迫(LD)、盐分胁迫(WS)和水盐联合胁迫(SY)3个胁迫处理,以无水盐胁迫处理作为对照(CK),研究水盐胁迫对玉米叶、根水势及玉米茎木质部水力特性的影响。与CK相比,水盐胁迫下玉米根、叶水势均有明显降低,叶-土水势差从大到小的顺序依次为CK>LD处理>WS处理>SY处理;茎干的初始水力导度K和茎流速率明显降低,其变化趋势与叶-土系统水势差变化趋势一致,相应地茎木质部水力导度损失百分比显著增大;同时,玉米叶片的净光合速率、蒸腾速率、气孔导度呈CK>LD处理、WS处理>SY处理,株高和叶片长度均表现为CK>LD处理>WS处理>SY处理。水盐胁迫均会降低玉米水势,使植株体内渗透势平衡遭到破坏,导致玉米光合性能受到抑制,使茎木质部内产生空穴化和栓塞现象,且不同胁迫方式导致不同程度的栓塞。
水盐胁迫;水力导度;栓塞;茎流;水势;光合作用
0 引言
植物茎干随着生长会进化出许多导管(管道分子或管胞),这些导管是植株长距离输水的重要通路,能够把根系吸收的水分和养分高效运输到叶片,保证植物的水分供应平衡[1]。因此,维持植物根-茎-叶内水分运输的连续性对植物的生存、生长极其重要。根据Dixon[2]提出的内聚力-张力理论,植物体内的水分是自下而上逆重力传输的,木质部内水分在负压以下是以连续水柱的形式存在的,其水分子间巨大的内聚力能够使水柱承受来自叶片蒸腾产生的张力[3]。通常处于负压下的水柱呈亚稳定状态,这种负压的程度由植物高度、土壤水分状况、植物蒸腾速率和土壤-植物水力路径阻力等因素综合决定[4-6]。
当某些外部因素(如干旱、盐害、低温等)改变了植物的蒸腾需求或者土壤水分有效性时,木质部负压超过阈值,会使气泡由气体填充的导管(或管胞)从纹孔膜上的小孔进入充水导管,使连续水柱断裂,并逐渐扩张形成栓塞,阻碍了木质部液流向上运输。这种空穴化诱导形成的栓塞会导致木质部水力导度降低,进而引起叶片水势下降和气孔关闭,严重抑制了植物的光合作用和作物生长[1,3]。
近年来,植物木质部栓塞的研究已成为植物耐旱性研究的热点[7-8],但研究大多集中在木本植物木质部水力特征与栓塞恢复的关系研究[9-13],对玉米等木质部较脆弱的禾本科植物的研究较少。鉴于此,在温度、空气湿度等生长条件较易控制的人工气候室内进行了桶栽试验,通过测定玉米各项生理生态指标,分析水盐胁迫对茎木质部水力特性的影响,探究玉米木质部栓塞的形成机理,为深入理解作物对水盐胁迫的响应提供数据支撑。
1 材料与方法
1.1 试验设计
选用登海605玉米品种为试验材料,试验用土取自田间0~30 cm的表层土,风干过5 mm筛后按体积质量1.35 g/cm3装桶,每桶装干土质量为24 kg。亚克力桶的尺寸为长20 cm,宽20 cm,高50 cm。试验于2019年在中国农业科学院新乡综合试验基地的人工气候室内进行。人工气候室控制温度为35℃/25℃(昼/夜),每天光照14 h(06:00—20:00)。试验共设置干旱胁迫(LD)、盐分胁迫(WS)、水盐联合胁迫(SY)和对照(CK)4个处理,CK为清水灌溉,水分控制在田间持水率的70%~75%,干旱胁迫(LD)处理用清水灌溉,水分控制下限在田间持水率的50%~55%;盐分胁迫(WS)处理用物质的量浓度为50 mmol/L的NaCl溶液灌溉,水分控制下限在田间持水率(质量含水率)的70%~75%;水盐联合胁迫(SY)处理用物质的量浓度为50 mmol/L的NaCl溶液灌溉,水分控制下限在田间持水率(质量含水率)50%~55%。每个处理设10个重复,见表1。
试验开始前,将所有重复的土壤含水率统一调整至田持的75%,在开始处理后,再将土壤含水率控制在各处理组的目标含水率区间内[14-16]。所有处理在进入拔节期前1周开始进行水盐胁迫处理,在此之前均用1/2 Hoagland营养液进行灌溉,配方为Ca(NO3)2·4H2O 0.59 g/L、KNO30.253 g/L、KH2PO40.068 g/L、MgSO4·7H2O 0.347 g/L、微量元素0.5 mL/L,其中微量元素配方为H3BO32.86 mg/L、MnSO4·H2O或MnSO4·4H2O 1.55或2.13 mg/L、ZnSO4·7H2O 0.22 mg/L、CuSO4·5H2O 0.08 mg/L、CuSO4·4H2O或(NH4)6MoO24·4H2O 0.09或0.02 mg/L、Na2Fe·EDTA或FeSO4·7H2O+Na2EDTA 24或(20.57+27.89)mg/L。利用便携式pH仪测定所有营养液,用稀盐酸调整pH值至5.8~6.2。

表1 水分胁迫及盐分胁迫试验设计
1.2 测定项目
1.2.1 株高和叶长
采用游标卡尺和直尺分别测量株高(从地面到所有叶片自然伸展时最高处的距离)和叶长(从叶枕到叶尖的距离),分别在开始处理前和处理2周后测量。
1.2.2 叶片气体交换参数
利用便携式光合仪Li-6400测定叶片的光合速率、蒸腾速率和气孔导度。分别在处理后1周(处理前期)、处理后3周(处理后期)测定,测定时选择每株玉米的功能叶,从上往下第3片叶,每片叶测量时选取叶片的中上部,避开叶脉,每个处理设4个重复,每次测定于09:00—11:00进行。
1.2.3 水势
使用WP4C露点水势仪测定叶水势、根水势、土水势。分别在处理后1周(处理前期)、处理后3周(处理后期)测量叶水势、土水势,根水势在处理后期进行破坏性取样测量。叶水势测量时取上部完全展开叶片中部位置测定,取样在08:00—09:00之间进行,用自封袋保存并放入装有冰袋的泡沫保温箱内待用。根水势测量选取10~20 cm土层根系,取出根系后清理干净附着残余土壤。每个处理设置4个重复。
1.2.4 茎干茎流与木质部水力导度
采用DYNAGAGE包裹式植物茎流计测量玉米实时茎干茎流,待玉米进入拔节期后安装茎流传感器,安装部位选取茎干直径与传感器大小合适且没有疤痕或损伤处。茎流计数据采用DT80和CR1000数据采集器24 h自动记录,设置数据采集时间间隔为每30 min采集1次。
木质部水力导度及栓塞程度使用XYLEM-Plus木质部导水率与栓塞测量系统测定,因测量需破坏性取样,故在处理后第3周进行测定。首先,从根部开始向上截取20 cm左右的茎段并将其放入水中,然后,在水中再进一步截取带有茎节的茎段,以避免因破坏性取样导致的茎段压力变化对茎木质部产生的影响。
假设有一段茎木质部没有栓塞,导水率为(K(mmol/(s∙Mpa)),干旱或者盐分胁迫以后,其导水率由K下降到,导水率的下降比可以通过式(1)得到:
100(1/K),(1)
如果=0,即K=K',也就是没有导管被栓塞。如果=100%,则=0,则所有导管都被栓塞[17-18]。栓塞程度大小用水力导度损失百分比来表示。
1.3 数据处理
采用SPSS 20.0和Microsoft Excel 2016进行数据统计分析与作图。显著性检验采用LSD、Duncan法。
2 结果与分析
2.1 水盐胁迫对玉米叶、根水势和土水势的影响
水盐胁迫下桶栽玉米叶、根、土水势见图1。由图1可知,水分胁迫、盐分胁迫和水盐联合胁迫均导致了玉米叶水势、根水势和土水势的降低、且规律基本一致,表现为SY处理<WS处理<LD处理<CK。与CK相比,LD、WS和SY处理的叶水势分别下降15%、16%、35%,且差异显著(<0.05);根水势分别下降19%、95%、166%,且差异显著(<0.05);土水势分别下降70%、192%、234%,且差异显著(<0.05)。
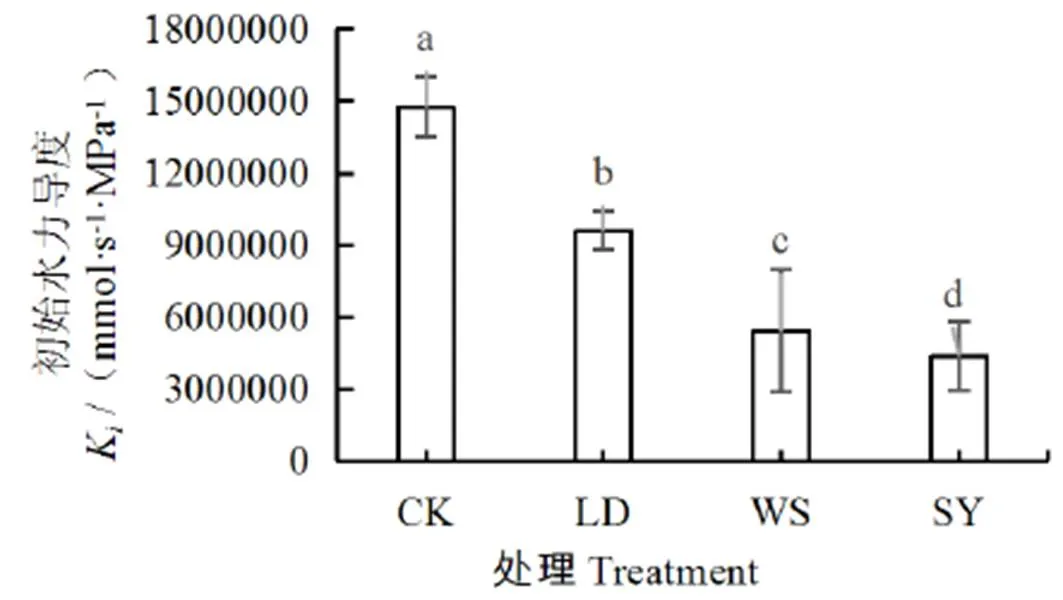
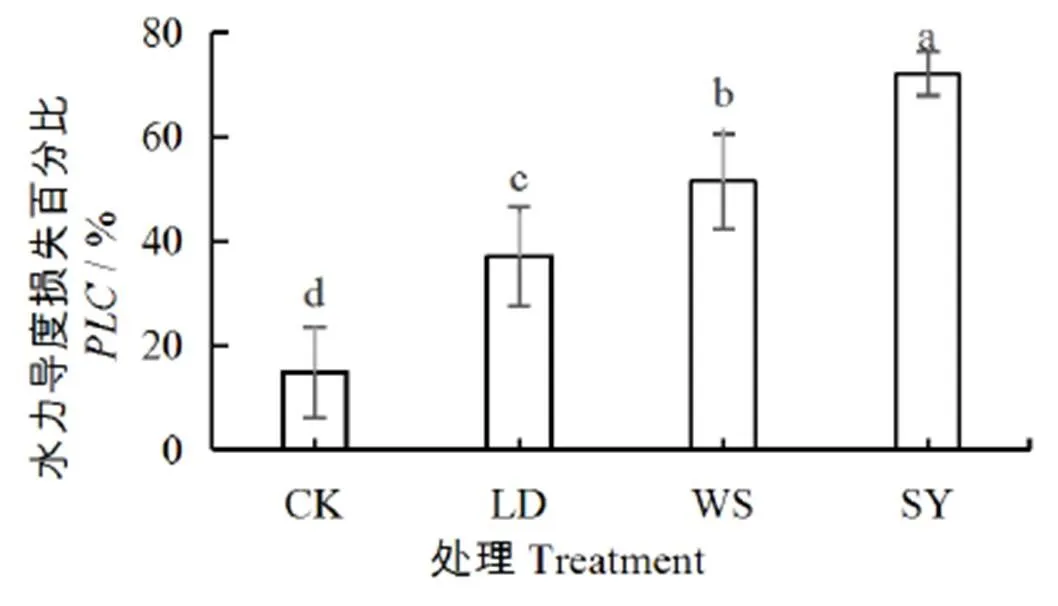
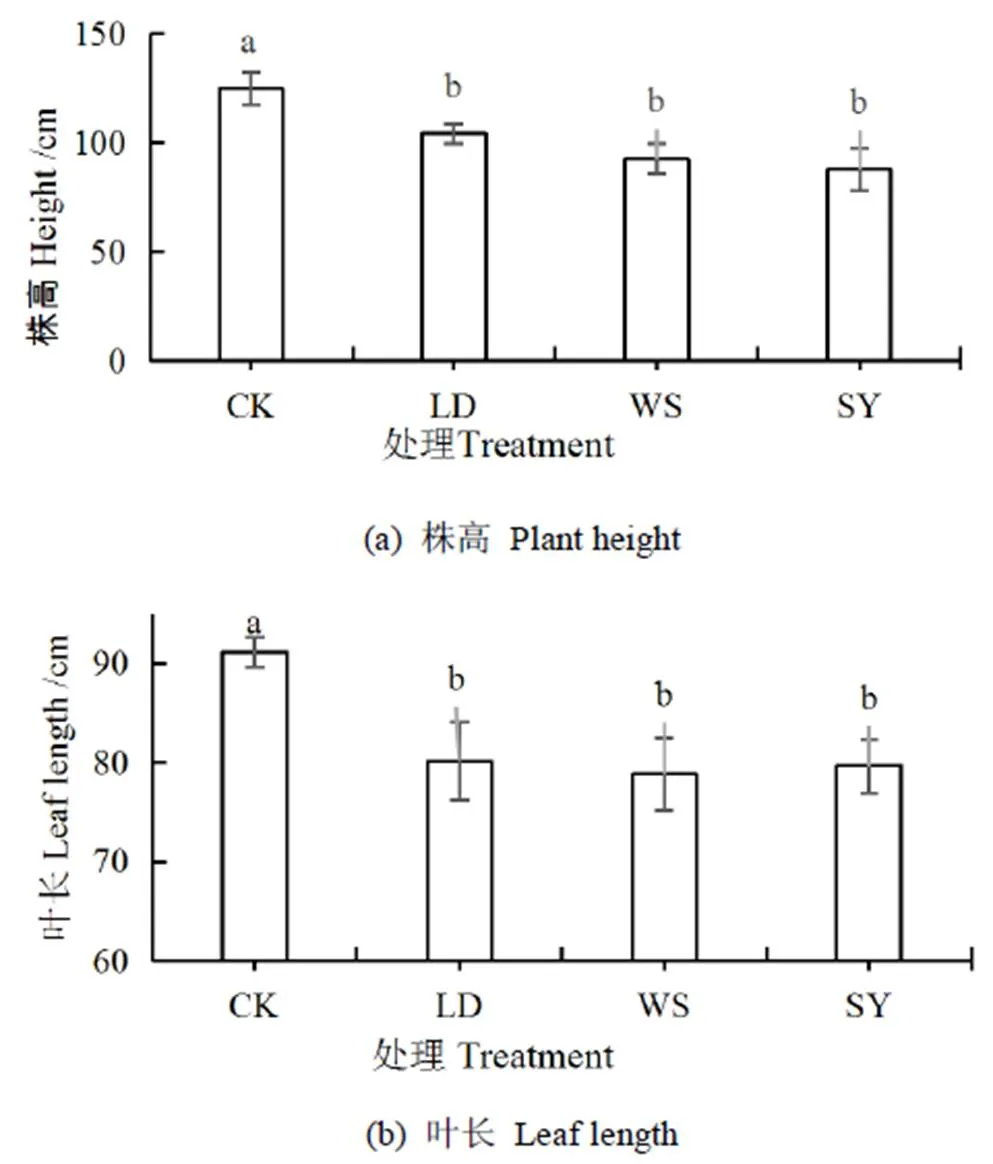
进一步分析得出,不同处理对桶栽玉米土壤作物系统的水势差影响显著,土-叶水势差从小到大依次为:SY处理 图1 水盐胁迫下玉米叶、根水势和土水势 茎干的初始水力导度K代表了植株茎干运输水分的能力,K越低,说明茎干输水能力越低。由图2可以看出,与CK相比,LD、WS和SY处理的K分别下降了35%、63%、71%,差异显著(<0.05),与根、土水势及土-叶水势差的变化趋势一致,即随着土壤水势降低,玉米茎干的初始水力导度K逐渐减小,即空穴化现象逐渐加重。 图2 不同处理下的玉米茎干初始水力导度 图3 不同处理下茎干的水力导度损失百分比PLC 水力导度损失百分比是表示木质部栓塞程度大小的重要参数。在图3中,与CK相比,LD、WS和SY处理的水力导度损失百分比分别增加了147%、243%、381%,即随着胁迫程度的增加而急剧增加,栓塞程度增高,差异显著(<0.05)。 图4 不同处理下玉米茎流速率日变化过程 图5 不同处理下玉米日茎流累积量变化 选择一个完整灌水处理周期(茎流计安装后第一次处理)各处理玉米茎流速率日变化过程进行分析,结果如图4所示。从玉米茎流速率日变化过程来看,每日的茎流变化都会经过一个“上升-峰值-下降”的完整循环,通常白天07:00左右各处理玉米的茎流开始快速上升,11:00左右茎流速率达到峰值,之后开始下降,21:00—次日06:00之间仅有微弱的茎流,降至最低值。由图4可见,CK、LD、WS和SY处理的茎流速率峰值分别是137.73、62.91、32.68、24.92 g/h,CK最大,胁迫条件下,茎流峰值随着胁迫程度增加而减小。 茎流日累计量与茎流速率密切相关。在图5中,不同处理下的茎流日累计量有显著差异,CK茎流日累计量最大,LD处理次之、WS和SY处理相近,显著低于CK和LD处理。 从图6可以看出,水分胁迫对叶片净光合速率、蒸腾速率、气孔导度均有显著的抑制作用(<0.05),盐分胁迫对叶片净光合速率等指标也有抑制作用,但下降幅度不及干旱胁迫,可能是由于干旱胁迫处理长期亏水导致的叶片损伤。水盐联合胁迫显著影响玉米的净光合速率和蒸腾速率(<0.05)。 图6 水盐胁迫下叶片气体交换参数 在非盐条件下,CK的净光合速率较干旱处理高49.2%,蒸腾速率与气孔导度分别高74.2%、65.8%,差异显著(<0.05)。在盐分处理下,盐分处理净光合速率较水盐联合胁迫高54.3%,蒸腾速率与气孔导度分别高出76.7%、71.8%,差异显著(<0.05)。 在同样水分梯度下,CK的净光合速率较盐分胁迫高10.6%,蒸腾速率与气孔导度分别高27.9%、18.1%,差异不显著(>0.05);干旱胁迫的净光合速率较水盐联合胁迫高19.6%,蒸腾速率与气孔导度分别高35.1%、32.5%,差异不显著(>0.05)。 叶片的光合性能越强,消耗的水分越多,对水分的需求就更迫切,对植株体内水分产生的蒸腾拉力越强,叶-根水势差也就应越大,图6各项气体交换参数中,CK>LD处理,WS处理>SY处理,总体保持高田持处理>低田持处理,无盐分处理>有盐分处理的趋势变化,这与水势和水力导度的变化趋势基本相同。 株高和叶长是直接反映作物生长状况的重要指标。由图7(a)、图7(b)和图1可以看出,随着胁迫程度增加,玉米植株高和叶长逐渐下降,与CK相比,LD、WS和SY处理的株高分别下降16%、26%、30%,叶长分别下降12%、13%、15%,各胁迫处理间较CK差异显著(<0.05),但各胁迫处理间无显著差异。 图7 水盐胁迫下玉米株高和叶长 干旱区盐碱地种植中,无论是干旱还是盐分对植物的生长都有抑制作用,并通过影响植物叶片光合特性指标和降低根茎水力特征指标等途径降低作物的产量和品质。当叶片的光合性能降低,影响到植物的代谢过程[19],就会导致水分亏缺,从而打破植株水分平衡,导致植株水势明显降低,并且随着胁迫程度的增强,下降幅度会增加[20]。在本试验中,CK的叶水势和根水势较3个胁迫处理高出了15%~166%,土壤-作物系统水势梯度的大小顺序依次为CK>LD处理>WS处理>SY处理,这与前人的研究结果一致。 当植株的水势下降后,植株茎木质部的张力就会增加,当这种张力增大到一定程度就会使植物体内的水柱断裂,从而形成空穴和栓塞[21-22]。所以,植物水势是与木质部栓塞关系最为密切的参数。在本试验中,玉米茎干的初始水力导度K在胁迫处理后逐渐减小,代表栓塞程度的水力导度损失百分比随着胁迫程度的增加,也急剧增加。相较于CK,3个胁迫处理分别增加147%~381%,栓塞程度从低到高为CK 作物水势下降是由于水盐胁迫导致作物的光合系统受到影响。任丽雯等[23]研究表明,长期严重的干旱(土壤相对湿度<42.7%)就会破坏玉米叶片的光合结构,导致光合速率和水分利用效率显著下降,严重影响作物正常生长,最终导致作物绝产;张春宵等[24]研究认为盐胁迫减少了玉米干物质的积累,造成了作物的减产,甚至绝产;李林等[25-26]研究表明,相较于正常浇水,干旱严重抑制花生的生长,使叶片失绿变黄叶绿素量下降,从而导致光合速率下降。因此,水盐胁迫会对叶片气孔运动和光合蒸腾产生不同程度影响。在本试验中,玉米植株生长及叶片光合性能在水盐胁迫下产生明显抑制,其中,干旱胁迫的各项气体交换参数数值低于盐分胁迫,异于水势与水力导度的变化趋势,这可能是由于长期干旱使叶片结构组织遭到受损而导致的,因为了避免灌溉后短时间内玉米水分充足,使各处理组短时间内各项参数无差异,所以每次测量都选在灌溉前1天,这时干旱处理组的玉米叶片已经出现了轻微萎蔫现象,导致各项气体交换参数数值偏低。但各处理间总体保持CK>LD处理,WS处理>SY处理的梯度趋势,具体为非盐条件下,CK净光合速率、蒸腾速率、气孔导度较LD处理高出49.2%~74.2%;在盐分胁迫下,WS处理较SY处理高出54.3%~71.8%。而在相同水分条件下,CK的各项气体交换参数较WS处理高出10.6%~27.9%,LD处理较SY处理高出19.6%~32.5%。综合来说,叶片光合性能在水盐胁迫下受到严重抑制,且水分胁迫比盐分胁迫造成的影响更为严重,这与任丽雯等[23]、李林等[25-26]的研究结果一致。 当水分胁迫下叶片气孔开度变小,蒸腾速率降低,使叶片出现不同程度的萎蔫现象,导致叶片的组织和细胞被破坏[27-28],且盐离子在细胞内大量积累,导致离子毒害和离子不平衡,较早看到的现象是叶片生长受到抑制[29]。在本试验中,CK的株高较3个胁迫处理组高出16%~30%,叶长高出12%~15%,作物生长在胁迫条件下受到显著抑制,生长指标从高到低依次为CK>LD处理>WS处理>SY处理,这与水势和水力导度的梯度的变化趋势相同,表明栓塞现象确实对作物生长确实有显著抑制作用。 1)水盐胁迫条件下,玉米的叶水势、根水势及土水势均降低,不同胁迫处理的叶-土水势差随着胁迫程度增加而减小,说明植株的吸水输水能力在胁迫条件下呈降低趋势。 2)当玉米受胁迫处理后,叶片各项气体交换参数受到影响,且叶片光合作用对茎木质部的蒸腾拉力在不同胁迫处理下会受到不同程度的抑制,植株整体的渗透势平衡遭到破坏,最终导致茎木质部出现空穴化现象和栓塞。 3)不同胁迫方式会导致不同程度的栓塞,栓塞程度越高,说明玉米茎干中的空穴化现象越严重,茎木质部输送水分就更加困难,茎流流速也就越低,体现在植株上的现象就是株高叶长减小,植株生长受到抑制。 [1] 金鹰, 王传宽, 周正虎. 木本植物木质部栓塞修复机制:研究进展与问题[J]. 植物生态学报, 2016, 40(8):834-846. JIN Ying, WANG Chuankuan, ZHOU Zhenghu. Mechanisms of xylem embolism repair in woody plants: Research progress and questions[J]. Chinese Journal of Plant Ecology, 2016,40(8):834-846. [2] DIXON DIXON H H, JOLY J. On the ascent of sap[J]. Proceedings of the Royal Society of London,1894(57): 3-5. [3] 陈志成, 姜丽娜, 冯锦霞,等. 木本植物木质部栓塞测定技术的争议与进展[J]. 林业科学, 2018, 54(5):143-151. CHEN Zhicheng, JIANG Lina, FENG Jinxia, et al. Progress and Controversy of Xylem Embolism Determination Techniques in Woody Plants[J]. Scientia Silvae Sinicae, 2018,54(5):143-151. [4] 万贤崇, 孟平.植物体内水分长距离运输的生理生态学机制[J]. 植物生态学报, 2007, 31(5):804-813. WAN Xianchong, MENG Ping. Physiological and ecological mechanisms of long-distance water transport in plants: A review of recent issues[J]. Journal of Plant Ecology, 2007, 31(5): 804-813. [5] KOCH G W, SILLETT S C, JENNINGS G M, et al. The limits to tree height[J]. Nature,2004,428(6 985): 851. [6] 刘畅, 陈志成, 刘晓静, 等. 光强和树体大小对锐齿栎树木水,碳平衡的影响[J]. 林业科学, 2017, 53(9):18-25. LIU Chang, CHEN Zhicheng, LIU Xiaojing, et al. Effects of Irradiation and Tree Size on the Carbon-Water Balances of Quercus aliena var. acuteserrata[J].Scientia Silvae Sinicae, 2017,53(9):18-25. [7] ZWIENIECKI M A, HOLBROOK N M. Confronting Maxwell's demon: biophysics of xylem embolism repair[J]. Trends in plant science,2009, 14(10): 530-534. [8] NARDINI A, GULLO M A L, SALLEO S. Refilling embolized xylem conduits: is it a matter of phloem unloading?[J]. Plant Science, 2011, 180(4): 604-611. [9] 安瑞, 孟凤, 尹鹏先, 等. 刺槐木质部栓塞脆弱性检测的方法比较[J]. 植物生态学报, 2019, 42(11):1 113-1 119. AN Rui, MENG Feng, YIN Pengxian, et al. Comparison of methods for detecting vulnerability of xylem embolism in Robinia pseudoacacia[J]. Chinese Journal of Plant Ecology, 2019,42(11):1 113-1 119. [10] 程向芬, 马晋, 赵涵, 等. 木本植物水力学结构之导管长度研究进展[J]. 植物生态学报, 2018, 42(6):609-618. CHENG Xiangfen, MA Jin, ZHAO Han, et al. Vessel length as a key hydraulic structure in woody plants: A review [J]. Chinese Journal of Plant Ecology, 2018, 42(6):609-618. [11] 于永畅. 旱柳皮层光合对木质部栓塞修复的驱动及其调节机制[D].北京:中国林业科学研究院, 2018. YU Yongchang. Repair and Regulation Mechanism of Xylem Embolism Driven by Cortex Photosynthetic in Salix Matsudana[D]. Beijing: Chinese Academy of Forestry, 2018. [12] 党维, 姜在民, 李荣, 等. 6个树种1年生枝木质部的水力特征及与栓塞修复能力的关系[J]. 林业科学, 2017, 53(3):49-59. DANG Wei, JIANG Zaimin, LI Rong, et al. Relationship between Hydraulic Traits and Refilling of Embolism in the Xylem of One-Year-Old Twigs of Six Tree Species[J]. Scientia Silvae Sinicae, 2017, 53(3):49-59. [13] WEI C. The transmission of gas pressure to xylem fluid pressure when plants are inside a pressure bomb[J]. Journal of Experimental Botany, 2000, 51(343):309-316. [14] 巩文军. 灌水方式对华北不同品种玉米产量形成和水分利用的影响[J]. 灌溉排水学报, 2018, 37(3):18-22. GONG Wenjun. Effect of Irrigation Patterns on Yield Formation and Water Use Across Various Maize Varieties in the North China Plain [J]. Journal of Irrigation and Drainage, 2018, 37(3):18-22. [15] 曹勇, 张建丰, 李涛, 等. 不同控光条件对春玉米耗水规律的影响[J]. 灌溉排水学报, 2018, 37(12):10-18. CAO Yong, ZHANG Jianfeng, LI Tao , et al. Effects of Light Intensity on Water Consumption of Spring Maize[J]. Journal of Irrigation and Drainage, 2018,37(12):10-18. [16] 孙成凡, 张志亮, 郑彩霞, 等. 间作模式下玉米干旱胁迫响应研究[J]. 灌溉排水学报, 2019, 38(S1):32-35. SUN Chengfan, ZHANG Zhiliang, ZHENG Caixia, et al. Response of Maize to Drought Stress under Intercropping Mode[J]. Journal of Irrigation and Drainage, 2019,38(S1):32-35. [17] STEPPE K. Diurnal dynamics of water flow through trees: design and validation of a mathematical flow and storage model, Ghent University[J]. Faculty of Bioscience Engineering, Ghent, Belgium, 2004. [18] MELCHER P J, MICHELE HOLBROOK N, BURNS M J, et al. Measurements of stem xylem hydraulic conductivity in the laboratory and field[J]. Methods in Ecology and Evolution, 2012, 3(4): 685-694. [19] 谭永芹, 柏新富, 朱建军. 等渗盐分与水分胁迫对三角叶滨藜和玉米光合作用的影响[J]. 生态学杂志, 2010, 29(5):881-886. TAN Yongqin, BAI Xinfu, ZHU Xinjian. Effects of isotonic NaCl and mannitol stress on the photosynthesis of Atriplex triangularis and Zea mays[J]. Chinese Journal of Ecology, 2010, 29(5):881-886. [20] 张洪旭, 杨德光, 李士龙, 等. 水分胁迫对玉米叶片水分代谢的影响[J]. 玉米科学, 2008(2):88-90. ZHANG Hongxu, YANG Deguang, LI Shilong, et al. Effects Under Water Stress on Leaf Water Metabolism in Maize[J].Journal of Maize Sciences, 2008(2):88-90. [21] 安锋, 张硕新, 赵平娟. 8种木本植物木质部栓塞变化与生理生态指标关系的研究(Ⅰ):与植物木质部水势的关系[J]. 西北植物学报, 2005, 25(8):1 595-1 600. AN Feng, ZHANG Shuoxin, ZHAO Pingjuan. Relations between Xylem Embolisms and Physiological Indexes in Eight Woody PlantsⅠ. Relationships with Xylem Water Potentials[J]. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(8):1 595-1 600. [22] 安锋, 蔡靖, 姜在民, 等. 8种木本植物木质部栓塞变化与生理生态指标间关系的研究 (Ⅱ)—与光合生理生态指标的关系[J]. 西北林学院学报, 2006, 21(1):37-42. AN Feng, CAI Jing, JIANG Zaimin, et al. Relationships between Xylem Embolism and Ecophysiological Index es in Eight Woody Plants in situ (II): The Relatio nship with Photosynthetic Ecophysiological Indexes[J]. Journal of Northwest Forestry University, 2006, 21(1): 37-42. [23] 任丽雯, 王兴涛, 赵鸿, 等. 春玉米叶片光合生理参数对土壤水分的阈值响应及其生产力分级[J]. 干旱区研究, 2019, 36(4):901-908. REN Liwen, WANG Xingtao, ZHAO Hong, et al. Threshold Responses of Leaf Photosynthetic Physiological Parameters of Spring Maize to Soil Moisture and Its Productivity Classification[J]. Arid Zone Research, 2019, 36(4): 901-908. [24] 张春宵, 刘文国, 袁英, 等. 玉米杂交种苗期耐盐碱筛选与大田鉴定的比较分析[J]. 玉米科学, 2010, 18(5):14-18. ZHANG Chunxiao, LIU Wenguo, YUAN Ying, et al. Comparative Analysis Between Salt-alkali Tolerance in Seedling Stage and Production in the Field of Maize Hybrid[J]. Journal of Maize Sciences, 2010, 18(5): 14-18. [25] 李林, 邹冬生, 刘登望, 等. 基于产量的花生基因型耐湿涝性综合评价[J]. 中国油料作物学报, 2004, 26(4):27-33. LI Lin, ZOU Dongsheng, LIU Dengwang, et al. Yield-based comprehensive evaluation on difference of waterlogging tolerance in peanut genotypes[J]. Chinese Journal of Oil Crop Sciences, 2004, 26(4):27-33. [26] 李林, 刘登望, 熊璟, 等. 花生生育早期耐涝性室内鉴定对大田期的意义[J]. 作物学报, 2008(3):477-485. LI Lin, LIU Dengwang, XIONG Jing, et al. Importance of Identification of Waterlogging Tolerance at Early Growing Stages of Peanut in Laboratory to Field Screening[J]. Acta Agronomica Sinica, 2008(3):477-485. [27] 袁国富, 庄伟, 罗毅. 冬小麦叶片气孔导度模型水分响应函数的参数化[J]. 植物生态学报, 2012, 36(5):463-470. YUAN Guofu, ZHUANG Wei, LUO Yi. Parameterization of water response functions in leaf stomatal conductance model for winter wheat[J]. Chinese Journal of Plant Ecology, 2012, 36(5): 463-470. [28] 刘战东, 肖俊夫, 刘祖贵, 等. 膜下滴灌不同灌水处理对玉米形态、耗水量及产量的影响[J]. 灌溉排水学报, 2011, 30(3):60-64. LIU Zhandong, XIAO Junfu, LIU Zugui, et al. Effects of Different Irrigation Treatment on Morphological Indexes, Water Consumption and Yield of Maize under Mulch Drip Irrigation[J]. Journal of Irrigation and Drainage, 2011, 30(3): 60-64. [29] 薛盈文, 王玉凤, 杨克军, 等. 外源NO对NaCl胁迫下玉米幼苗生长和渗透调节能力的影响[J].干旱地区农业研究, 2016,34(2):171-176, 200. XUE Yingwen, WANG Yufeng, YANG Kejun, et al. Effects of exogenous nitric oxide on growth and osmoregulatory capability of maize (Zea mays L.) seedlings under salt stress[J]. Agricultural Research in the Arid Areas,2016,34(2):171-176, 200. Change in Hydraulic Characteristics of Maize in Response to Water and Salinity Stresses XU Jianxin1, WANG Qian1, GAO Yang2*, SUN Jingsheng2, HOU Zelin1 (1. North China University of Water Resources and Electric Power, Zhengzhou 450000, China; 2. Farmland Irrigation Research Institute, Chinese Academy of Agricultural Sciences, Xinxiang 453002, China) 【】Drought and salinity are most common abiotic stresses impending crop growth, and understanding how crops hydraulically respond to these stresses is imperative for improving agronomic management. In this paper, we present the results of an experimental study on the impact of water and salinity stresses on water characteristics of maize.【】The experiment was conducted in a phytotron and consisted of four treatments: water stress (LD), salt stress (WS), coupled water and salt stress (SY); the control was without stress (CK). In each treatment, we measured leaf water potential, root water potential and stem hydraulic conductivity of the maize. 【】Compared with CK, LD, WS and SY reduced water potential in both root and leaf noticeably. In all treatments, the difference between water potential in the leaf and the soil was in the order of CK>LD>WS>SY. Water and salinity stresses both reduced initial hydraulic conductivity and sap flow rate in a similar way as their impact on the difference between water potential in the leaf and the soil; they led to an increase in the percentage of hydraulic conductivity loss of the stem xylem (PLC). Water stress reduced net photosynthetic rate, transpiration rate and stomatal conductance, and coupling water and salt stresses reduced these traits further. Plant height and leaf length in all treatments was in the order CK>LD>WS>SY.【】Water and salt stresses in separation or in coupling both reduced water potential and imbalanced osmotic potential in the plant, thereby inhibiting photosynthesis and embolizing the stem xylem network, although the embolization varied with types and degree of the stresses. water and salt stress; hydraulic conductivity; embolism; sap flow; water potential; photosynthesis A 10.13522/j.cnki.ggps.2019300 1672 - 3317(2020)01 - 0045 - 07 徐建新, 王潜, 高阳, 等. 水盐胁迫对玉米茎木质部水力特性的影响[J].灌溉排水学报, 2020, 39(1):45-51. XU Jianxin, WANG Qian, GAO Yang, et al. Change in hydraulic characteristics of maize in response to water and salinity stresses [J]. Journal of Irrigation and Drainage, 2020, 39(1): 45-51. 2019-10-03 国家自然科学基金重大项目(51790534);国家自然科学基金项目(51879267) 徐建新(1954-),男。教授,博士生导师,主要从事区域水资源高效利用方面的研究。E-mail:xujianxin@ncwu.edu.cn 高阳(1978-),男。研究员,主要从事作物水分生理与高效用水研究。E-mail:gaoyang@caas.cn 责任编辑:白芳芳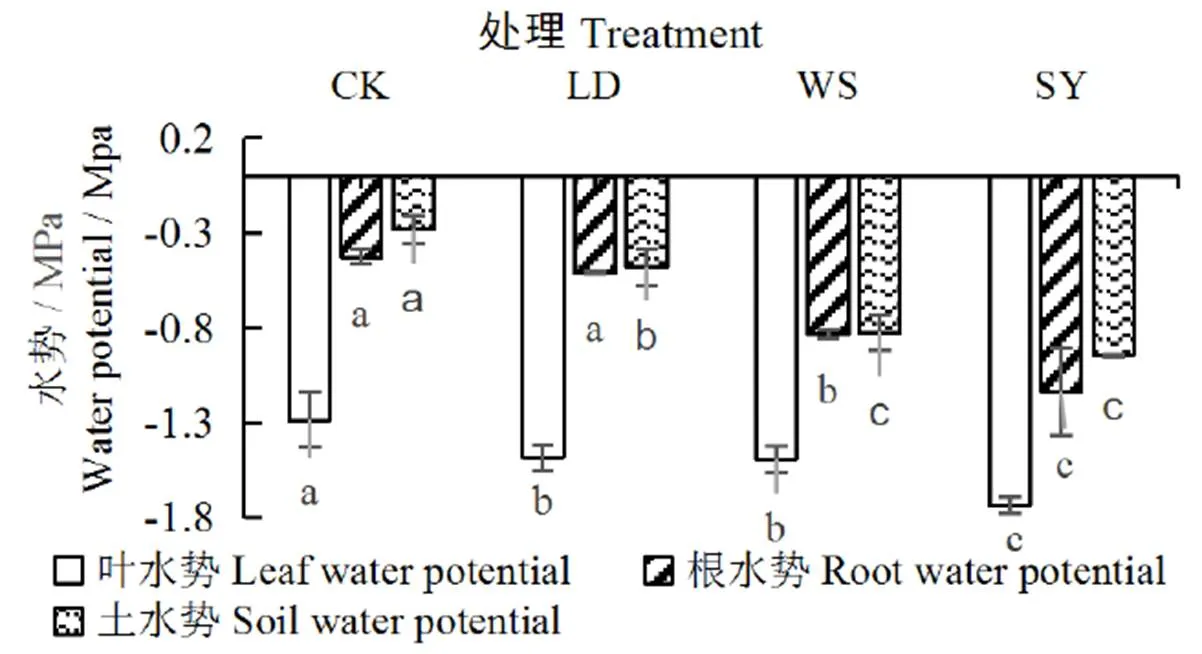
2.2 水盐胁迫对玉米茎木质部水力导度与水力导度损失百分比的影响
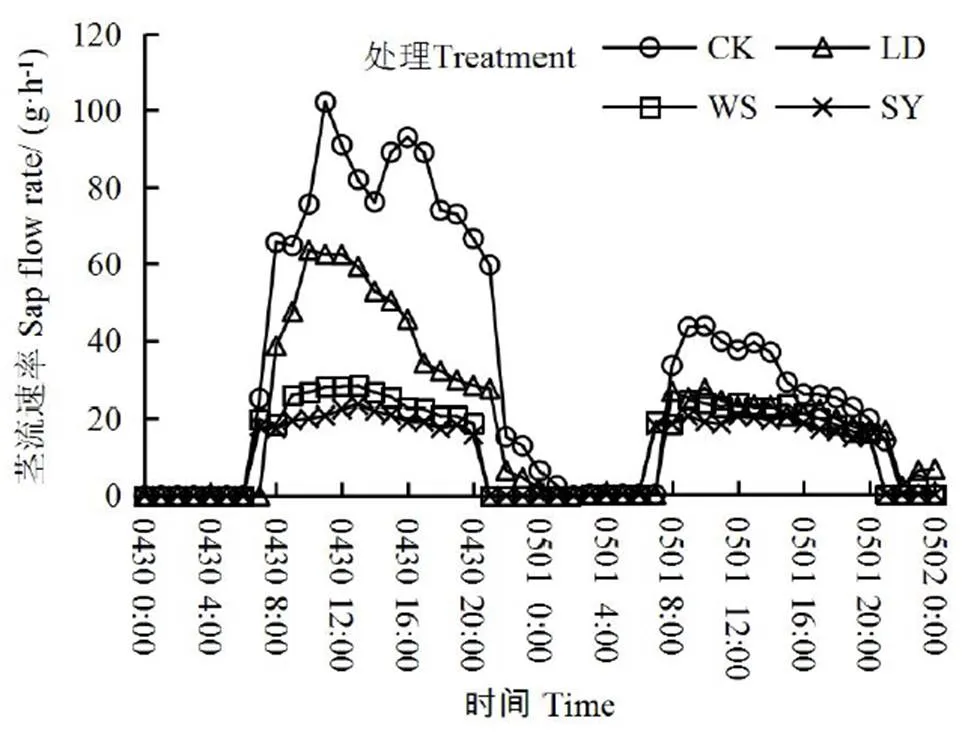

2.3 水盐胁迫对玉米茎木质部茎流的影响
2.4 水盐胁迫对叶片气体交换参数的影响

2.5 水盐胁迫对玉米生长指标的影响
3 讨论
4 结论
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
海洋湖沼通报(2022年3期)2022-06-29 01:23:22
科学技术与工程(2021年22期)2021-09-09 05:57:44
无机盐工业(2021年8期)2021-08-12 07:21:26
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
四川地质学报(2020年2期)2020-05-31 06:52:56
水利科技与经济(2016年3期)2016-04-09 13:57:20
水利科技与经济(2016年4期)2016-04-09 03:49:18
