
耐盐细菌研究进展及多样性分析
2020-03-15 09:55詹建立易霞
高师理科学刊 2020年12期
詹建立,易霞
(1.喀什大学 生命与地理科学学院,新疆 喀什 844000;2.九江学院 基础医学院,江西 九江 332005)
耐盐细菌(B.halodurans)是指对2.5 mol/L 以上浓度氯化钠有特殊适应能力的细菌.它主要生活在盐湖、盐碱湖、海水、废水、盐碱地、活性污泥、腌制食品中.由于长期生活在极端环境下,菌体细胞内形成多种具有特殊功能的酶以适应环境,进而在物种、基因和生理代谢等方面存在多样性.
国外对耐盐菌的研究起步较早.Arreola R[1]等对耐盐芽孢杆菌的转录因子PyrR 进行了初步研究;Anbu Rajan L[2]等对四氢嘧啶的生物合成基因的进化关系和异源表达进行了研究;杨重[3]等从基因层面对2种耐铬盐还原菌Staphylococcus sp.YZ-1 和Bacillus cereusCC-1 的Cr(Ⅵ)脱毒特性与机理差异进行了研究;曹凯[4]等对浙江舟山盐场来源的耐盐菌(Bacillussp.CK-1)蛋白酶的酶学特性进行了研究,结果显示该菌分泌的蛋白酶活性随着NaCl 浓度的变化呈现先增大后减小的趋势.本文主要对B.haloduransC-125,MIR32,DSM497,XJU-1,XJU-80 等亚种在物种进化、代谢生理、遗传和生物多样性等方面进行归纳,并分析该属种存在种间多样性的可能原因.
耐盐菌研究主要集中在菌株的分离鉴定、功能菌株的开发利用、酶学、有机物的降解和去除、发酵性能、植物促生特性、有毒物质的脱毒机理等领域.耐盐菌作为极端微生物,在细胞内形成多种具有特殊功能的酶以适应环境.因此,酶资源的研究和开发一直是该菌株研究的热点.Nquyen T T[5]等探讨了耐盐芽孢杆菌Bh0493 的催化功能;耐盐芽孢杆菌C-125 的Bh0225 的一种未知功能的酶编码基因已经成功克隆.Hong Y H[6]等对耐盐芽孢杆菌AI 品系依赖于金属的超嗜热的L-阿拉伯糖异构酶进行了同源重组.Agarwal R[7]等还对L-核酮糖激酶的底物多样性进行了初步思考.木聚糖酶、漆酶和角蛋白酶的研究也有开展[8-10].当然,耐盐芽孢杆菌的耐盐机制仍需深入探讨.
2 耐盐细菌研究进展
我国学者在功能菌株的开发利用方面也有大量研究.闫红梅[11]从环氧树脂废水中分离得到产碱菌、假单胞菌、芽孢杆菌3 株耐盐菌株,对其降解特性进行了研究;宋郭柳[12]等利用菌株分离、电镜观察和DNA扩增技术从深圳填海区植物根际土壤中筛选鉴定了3 株耐盐菌,并对其碳利用能力进行了分析;陈远哲[13]等研究了金华火腿来源的耐盐菌(Staphylococcus equorum)发酵对咸鸭蛋蛋清脱盐及抗氧化活性的影响;孙晓莹[14]等对盐碱地植物根际土壤来源的耐盐菌(Pseudomonas brassicacearumYZX4)的植物促生特性进行了研究;陈弘昊[15]等对3 株耐盐细菌(S1Microbacterium sp,G12Zhihengliuella sp,Y3Pseudomon putida)对多环芳烃降解性能进行了研究.
2.1 菌株分离鉴定
迄今为止,已分离鉴定的B.halodurans有很多,典型种如B.haloduransC-125(JCM9153,AB002661),MIR32(AY017348),DSM8718(X76442),DSM497(AJ302709),AH-101(JCM9161,AB027713),XJU-1(CGMCC 1.4010,AY856453),XJU-80(CGMCC 1.4011,AY856452)等.B.haloduransC-125 分离于1977年[16],Horikoshi 先后报道了其分泌产生的β-半乳糖苷酶和木聚糖酶[17-18].B.haloduransC-125(JCM9153,AB002661)是继B.subtilis之后的又一株全基因测序的Bacillus,目前已经推测其有4 066 个编码蛋白.MIR32(AY017348)于1995 年由Breccia 分离于阿根廷[19].2002 年,M Alejandra Martinez[20]等通过16S rRNA 序列分析、16S-23S rRNA PCR 指纹、tDNA 间区分析和质粒转化实验将MIR32 鉴定为B.halodurans.DSM8718于1993 年分离于德国,于1994 年初步将其鉴定为B.halodurans[21].AH-101(JCM9161,AB027713)由Takami 于1989 年分离于日本,最初将其鉴定为能够产耐热角蛋白酶的芽孢杆菌(Bacillus)[22].1999 年Takami等又进一步将AH-101 鉴定为B.halodurans[23].DSM497(AJ302709)于2000 年由Swiderski 分离于德国.B.haloduransXJU-1 和B.haloduransXJU-80 是国内报道的又一株耐盐芽孢杆菌[24],与南美(阿根廷)、亚洲(日本)和欧洲(俄罗斯)等地分离到的B.halodurans种进行了研究.B.haloduransAlk36 是一株嗜碱(pH7.5)耐热的G+细菌,其嘌呤核苷磷酸化酶也得以克隆和纯化[25-26].这些研究对阐明B.halodurans属内种的遗传多样性具有重要意义.
2.2 多样性研究
2.2.1 物种多样性B.haloduran有很多亚种,如C-125,MIR32,AH-101,DSM497,DSM8718,ALK36.这些种在地理分布上存在差异.XJU-1 和XJU-80 同是分离于中国吐鲁番水泥厂的污水中,但是基于16S rRNA序列分析不在同一分支上,前者与分离于日本的C-125 在同一分支上,后者与分离于德国的DSM497在同一分支上.同时是分离自日本的C-125 和AH-101 不在同一分支上,同是分离于德国DSM8718和DSM497也不在同一分支上.可以推断同属一物种即便是在同一自然环境下,也可以演化为不同的亚种.相同因素胁迫下,微生物的进化也是多样性的(见图1).
2.2.2 生理代谢多样性B.haloduransC-125,MIR32,AH-101,XJU-80,XJU-1 在形态和生理生化方面表现出属内的诸多共性,如G+、棒状细胞、孢子大多椭圆且多膨胀、好氧生长、水解淀粉、液化凝胶.但是这些菌株在表型上呈现出生理和代谢的多样性(见表1).如细胞大小不一,孢子形状和膨胀部分不同(如卵形和轻微膨胀),生长的温度和pH 范围有差异,NaCl 耐受程度不同,吐温降解有差异.这些生理和代谢的多样性很可能源于菌种生活环境的不同.因为不同生活环境给微生物细胞的胁迫不同,为了适应环境,同一种的细胞可能会进行不同的代谢,进而有了不同的产物.
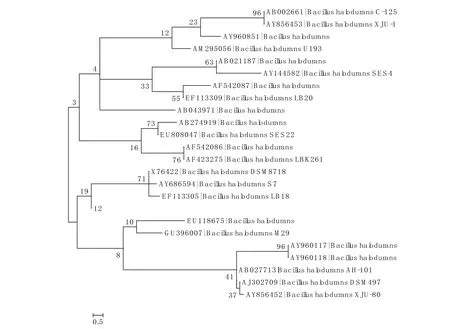
图1 B.halodurans 菌株进化树

表1 B.halodurans 菌株的形态和生理生化特征
2.2.3 基因多样性(1)16S rRNA 序列.16S rRNA 基因的很多特点有利于进行生物进化和系统分类研究,如其普遍存在于原核生物细胞,信息恒定,含量较高且易提取,分子量适中.因此,在B.haloduran种的分类也采用16S rRNA 基因.图1 说明B.haloduran种内各菌株间分子进化关系的同时,也存在16S rRNA序列的多样性.分离于新疆吐鲁番的菌株XJU-1 和XJU-80 在进化上并不在同一分支上,而与分离自日本的C-125 和分离自德国的DSM497处于同一分支上,这说明相同生境下的菌株存在基因序列的多样性.
(2)CG 含量分析(见表2).CG 含量是目前发现微生物新种所必需的指标之一.B.haloduransCG含量的平均值为43.7%,44.5%,44.1%,42.2%,40.5%,43%.所选6 种菌株(C-125,AH-101),DSM497,(XJU-1,XJU-80),MIR32 分别分离于日本、德国、中国,阿根廷,在地理环境上有显著的差异,在物种进化和遗传多样性方面也有代表性.部分B.halodurans菌株的CG 含量不在此范围,如DSM49(744.5 mol%)和XJU-80(40.5 mol%).这就体现出B.halodurans在种上的多样性.据测定CG 比相差低于2%时,没有分类学上的意义;种内各菌株间的(C+C)mol%含量差别可在2.5%~4.0%之间,若相差在5%以上时,就可以认为属于不同种,假如差距超过10%,一般就认为是不同属.

表2 B.halodurans 菌株16S rRNA 和CG 含量
(3)DNA-DNA 同源性杂交(见表3).物种DNA 碱基的排列顺序是长期进化的历史在分子水平上的纪录,DNA-DNA 同源性杂交是更精确的遗传性状指标.亲缘关系越近的微生物,其碱基序列也越接近,反之亦然.一般认为,若两菌间CG 比相差1%,则碱基序列的共同区域就约减少9%;若CG 比相差10%以上,则两者的共同序列就极少[12]62-67.若有一群CG 比范围在5%以内的菌株,要鉴定它们是否属同一个物种,就必须通过DNA-DNA 间的分子杂交.

表3 B.halodurans 菌株DNA-DNA 同源性杂交
由表2 和表3 可见,CG 比范围在5%以内的C-125,XJU-1,XJU-80,DSM497菌株,在DNA-DNA 同源性杂交结果上显示明显的差异性.
3 讨论
3.1 多方面综合考虑微生物谱系分析
在生态环境中,由于环境因子的组成和影响条件不同,各个生境中的微生物类群的分布和组成就有所不同,B.halodurans物种的多样性印证了这一点.即便是在相同的生境中,同一物种下不同亚种的16S rRNA序列分析也会有很大的差异,如XJU-1,XJU-80 同是分离于中国吐鲁番水泥厂的污水中,16S rRNA 序列分析却不在同一分支上.因此,对微生物的谱系分析应该从基因遗传、生理和生态等各方面综合考虑.
3.2 基因改变背景下的环境胁迫是种间多样性的原因
B.halodurans种内,不同亚种在形态和生理生化方面表现出诸多共同特征,列举的几种B.halodurans亚种在革兰氏反应、细胞形状、好氧生长、生长温度、水解淀粉、凝胶液化、乙酸利用、氧化酶、木聚糖方面差异很小(见表1).说明这几种微生物在细胞壁的结构、对温度的适应、代谢类型、次生产物等方面有共性,它们可能存在乙醛酸途径来获取能量和营养.因此,环境胁迫对于生理代谢的影响和基因相比显得更为直接,但遗传基因改变才是环境影响的背景因素,也是种间多样性的可能原因.
4 展望
作为极端微生物,耐盐菌研究主要集中在菌株的分离鉴定,功能菌株的开发利用、酶学、有机物的降解和去除、发酵性能、植物促生特性、有毒物质的脱毒机理等方面.利用极端环境胁迫下耐盐菌不同亚种在形态和生理生化方面表现出来的特性,进行功能菌株的筛选、改良,并建立功能菌株库,这应该是耐盐菌今后研究的主要趋势.随着经济的迅速发展,环境保护也愈显迫切.因此,能够分解有机物、净化污水的功能菌株应用会更加广泛.
猜你喜欢
热带亚热带植物学报(2022年4期)2022-08-18
昆虫学报(2021年11期)2021-12-08
今日农业(2021年21期)2021-11-26
——和田盘羊
野生动物学报(2021年4期)2021-11-14
湖南饲料(2021年4期)2021-10-13
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
食品与发酵工业(2019年18期)2019-10-11
天然产物研究与开发(2018年9期)2018-10-08
