
云南省剑川县鼠疫疫源地分离菌株基因组多态性
2020-03-13 11:51:08石丽媛丁奕博张海鹏段存娟谭红丽董珊珊钟佑宏
中国人兽共患病学报 2020年1期
石丽媛,丁奕博,张海鹏,郭 英,段存娟,谭红丽,董珊珊,钟佑宏,王 鹏
剑川县隶属云南省大理白族自治州,东邻鹤庆县,南接洱源县,西与兰坪县、云龙县接壤,北与丽江市毗连。县境东西横距58 km,南北纵长55 km,面积约2 250 km2。剑川县记载的鼠疫流行史最早可以追溯到1898年金华镇一带发生鼠疫流行[1];1950年甸南镇有流行;1954年沙溪镇大长乐村出现多例腺鼠疫病例;1975年从沙溪镇石龙村1只自毙小林姬鼠分离到鼠疫菌,证实了剑川县存在野鼠鼠疫疫源地[2]。该疫源地1975—1990年期间流行较为活跃,每年都有动物间鼠疫流行[3],但未发现人间病例。1998年至今,该疫源地处于相对静息期,未监测到动物鼠疫流行信息。2017年4月从金华镇庆华村委大庆村居民区捡获1只自毙黄胸鼠,从鼠及其寄生棕形额蚤各分离到1株鼠疫菌[4]。这是剑川县鼠间流行静息20年后,从距离原疫点-石龙村约25 km的地方发现的新的动物间鼠疫。
上世纪50年代分离菌株与70年代后分离菌株有何关系,2017年新疫点分离菌株与之前流行的菌株之间又是什么关系,弄清这些问题,对于全面认识剑川疫源地的性质有着重要意义。本研究对剑川县几十年累积保存的鼠疫菌株进行基因组多态性研究,以阐明该疫源地鼠疫菌的种群演化及与毗邻疫源地菌株之间的遗传进化关系。
1 材料与方法
1.1菌株来源及其DNA制备 按时间分布随机选取剑川县境内分离菌株,兼顾不同疫点、不同分离源,共计24株,菌株具体信息见表1。同时选择其他鼠疫菌株26株作为对照:包括云南家鼠疫源地保山、德宏、普洱、临沧、版纳、红河、文山菌株各1株,云南毗邻的贵州、广西家鼠疫源地菌株各2株及丽江野鼠疫源地菌株15株。菌株DNA提取制备采用Qiagen DNeasy Blood & Tissue Kit 试剂盒进行。EV76作为PCR反应的阳性对照。
1.2差异区段(Different region,DFR)分型 包括23对DFR分型引物(DFR01-23)和质粒验证引物(PMT1)[5]。扩增条件:95 ℃ 3 min;94 ℃ 30 s,60 ℃(DFR18、DFR19用56 ℃)30 s,72 ℃ 1 min,30个循环;72 ℃ 5 min。琼脂糖凝胶电泳检测PCR产物。
1.3规律成簇的间隔短回文重复序列(Clustered regularly interspaced short palindromic repeats, CRISPRs)分型 共有YPa、YPb及YPc 3对引物[6]。扩增条件:95 ℃ 5 min;95 ℃ 40 s,58 ℃ 40 s,72 ℃ 40 s,30个循环;72 ℃ 5 min。将产物测序,利用在线工具(http://crispr.i2bc.paris-saclay.fr/)查找spacers[7],再对应Spacers库识别出spacer的类型。
表1 24株剑川县境内分离菌株信息表
Tab.1 Information of 24 strains isolated from Jianchuan County

分离年 疫点信息数量 分离源1954剑川县沙溪镇长乐村大长乐3大绒鼠1,黄胸鼠1,病人11957剑川县甸南镇狮河村下河1绒鼠怪蚤1975剑川县沙溪镇石龙村1小林姬鼠1977剑川县沙溪镇长乐村鹅颈山1方叶栉眼蚤1979剑川县羊岑乡新松村1大足鼠1979剑川县沙溪镇石龙村1棕形额蚤1980剑川县沙溪镇石龙村1特新蚤指名亚种1982剑川县沙溪镇石龙村1齐氏姬鼠1983剑川县沙溪镇石龙村2大绒鼠1,大足鼠11984剑川县沙溪镇石龙村4齐氏姬鼠1,特新蚤指名亚种1,锐额狭臀蚤1,黄胸鼠11985剑川县沙溪镇石龙村2齐氏姬鼠1,方叶栉眼蚤11989剑川县沙溪镇马坪关2齐氏姬鼠1,大绒鼠11990剑川县沙溪镇石龙村1大耳林姬鼠2017剑川县金华镇庆华村大庆3特新蚤指名亚种1,棕形额蚤1,黄胸鼠1
注:分离源后的数字代表菌株数量。
1.4多位点可变数目串联重复序列 (Multiple-locus variable -number tandem repeat analysis,MLVA)分型 采用MLVA“14+12”分型策略[8]的26个可变数目串联重复序列(Variable Number of Tandem Repeats,VNTR )位点。扩增条件:95 ℃ 5 min;95 ℃ 1 min, 52/55/60 ℃ 1min(不同引物退火温度不同),72 ℃ 1min,30个循环;72 ℃ 5 min。将产物进行毛细管电泳检测大小,将产物碱基数与上下游序列碱基数的差值除以核心序列的长度算出重复数。
1.5聚类分析 将上述3种方法的数据导入BioNumerics 6.6。将50株实验鼠疫菌株及2株外围对照株(0.PE 布氏田鼠鼠疫菌91001、L1970015)的DFR和CRISPRs数据(在“Experiments”中同时选中“DFR”和“CRISPRs”),运行“Clustering”下的“Advanced cluster analysis”下的“MST for categorical data ”生成最小生成树(Minimum spanning tree analysis,MST)。再将基于DFR+CRISPRs的MST中的簇进行MLVA26分型,再次生成该簇的MST。
2 结 果
2.1基于DFR+CRISPRs的MST 50株实验鼠疫菌株被分成3个簇和1个独立株(剑川菌株YNJC20),如图1所示。24株剑川疫源地菌株中有20株菌聚为一个簇(红色底色),称为剑川野鼠鼠疫簇;15株丽江野鼠疫源地菌株全部聚为一个簇(绿色底色),称为丽江野鼠鼠疫簇,剑川2017年疫情分离3株菌位于该簇中;11株家鼠鼠疫疫源地菌株全部聚为一个簇(蓝色底色),称为家鼠鼠疫簇。 与外围菌株0.PE关系最近的丽江野鼠鼠疫簇。
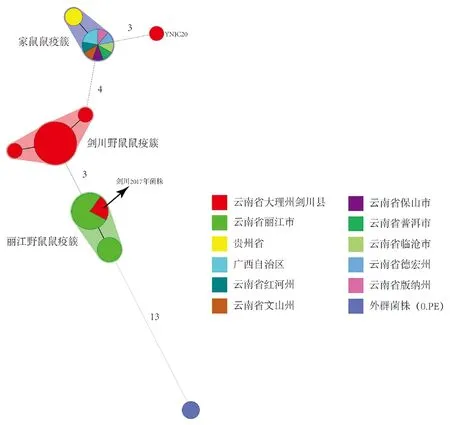
注:以菌株分离地点创建Group赋予不同颜色,圆的大小代表菌株数量,同一圆内的菌株具有相同的DFR&CRISPRs基因型,有相同底色的圆为一个簇,同一簇内的位点差异数不超过1。图1 50株实验鼠疫菌及2株0.PE菌株基于DFR和CRISPRs位点生成的MSTFig.1 Minimum spanning tree analysis of DFR & CRISPRs to 50 experimental strains and 2 0.PE strains
2.2剑川野鼠鼠疫簇基于MLVA26的MST 剑川野鼠鼠疫簇共20株菌,全部分离自大理州剑川县,分离时间为上个世纪50、70、80、90年代(按照不同分离年代赋予颜色)。如图2所示,整体而言,邻近菌株间位点差异不超过3,50年代菌株与其邻近的80年代菌株间位点差异也未超过3。
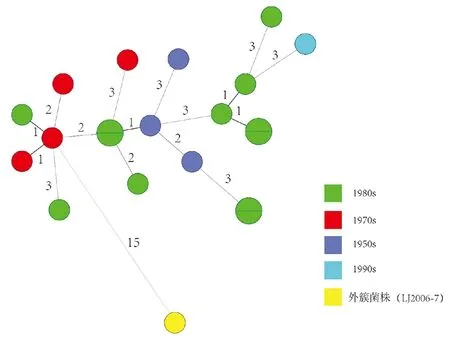
注:以菌株分离年代创建Group赋予不同颜色,同一圆内的菌株具有相同的MLVA基因型。图2 20株剑川野鼠鼠疫簇菌株基于MLVA26的MSTFig.2 Minimum spanning tree analysis of MLVA26 to 20 strains in Jianchuan Wild Rodents Plague Cluster
2.3剑川2017年分离菌株基于MLVA26的MST 剑川2017年菌株在DFR+CRISPRs分型中被划分到了丽江野鼠鼠疫簇中,利用MLAV26对其与丽江15株菌进行聚类分析如图3所示。从图中可以看出剑川2017年分离的3株菌株具有相同的MLVA基因型,与丽江2017年从玉龙县鹿子村分离的菌株(2017LZ)仅有2个VNTR位点的差异(N2577,M23)。
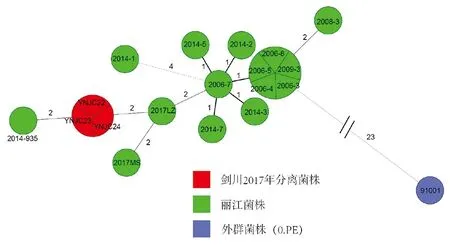
注:以菌株不同来源创建Group赋予不同颜色,同一圆内的菌株具有相同的MLVA基因型。图3 剑川2017年分离菌株与丽江野鼠鼠疫簇菌株基于MLVA26个位点生成的MSTFig.3 Minimum spanning tree analysis of MLVA26 to isolates from Jianchuan outbreak in 2017 and Lijiang Wild Rodents Plague Cluster
3 讨 论
细菌基因组多态性产生有着不同的遗传机制,基因水平转移和缺失、短重复序列、极性化记录噬菌体入侵事件等分别对应DFR、MLVA、CRISPRs这3种分型方法,从不同角度描述细菌基因组多态性[9]。云南鼠疫菌以往的分子分型研究[10-12]表明,DFR、MLVA、CRISPRs分型能区分菌株生态型,尤其是MLVA,其包含大量多态性信息,能够识别个体特征,是三者中分辨率最高的。本研究将3种分型方法整合,以DFR+CRISPRs两种分型数据先行大尺度分类,再以MLVA26进行细微遗传差别分析,阐释了剑川鼠疫菌在云南鼠疫菌中的分型位置及其种群的演化。
3.1剑川2017年新发动物鼠疫疫情是剑川县 新的一种鼠疫类型 剑川2017年新发疫情位于金华镇庆华村,与上世纪70—90年代剑川野鼠鼠疫好发疫点沙溪镇石龙村之间直线距离25.5 km。在DFR+CRISPRs分型中,剑川2017年菌株被划分到了丽江野鼠鼠疫簇,在后续与丽江菌株的MLVA聚类分析中,与丽江菌株仅有2个VNTR位点的差异。说明该次疫情并非上世纪70—90年代剑川野鼠鼠疫(石龙村为中心)的传播复燃,而是丽江野鼠鼠疫向南进一步扩散的结果。事实上,在剑川2017年疫情发生之时,丽江和大理州鹤庆也发现了鼠间疫情,且鹤庆分离菌株分子溯源上与丽江菌株高度同源,也仅有2个VNTR位点的差异[13]。因此认为,剑川2017年疫情是剑川县一种新的鼠疫类型。
3.2上世纪50年代的剑川鼠疫疫情具备剑川野鼠鼠疫遗传特征 剑川野鼠疫源地(滇西山地齐氏姬鼠、大绒鼠鼠疫自然疫源地)的发现源自1975年从剑川沙溪石龙一自死小林姬鼠体内分离出鼠疫菌。据既往文献记载,1954年剑川沙溪长乐村鼠疫流行中从患者、大绒鼠、黄胸鼠体内分离出的鼠疫菌为家鼠鼠疫菌株。本研究3种分型方法表明50年代剑川流行株与70年代以后的流行株在分子分型上是同一类型的菌株,即同为剑川野鼠鼠疫性质的菌株。据我所内部资料表明1954年剑川沙溪长乐村鼠疫流行中分离的3株菌在初次生化检测中发现甘油发酵-,麦芽糖+,与家鼠菌株生化特性一致,且由于同时期在大理州的其他4市/县(大理、祥云、弥渡、巍山)也发生了人间鼠疫,因此把50年代剑川菌株定性为家鼠鼠疫性质。但内部资料也表明,后来再次检测这些菌株发现甘油发酵+,麦芽糖+,王鹏等[14]对这些菌的检测结果也是甘油发酵+,麦芽糖发酵阳性/阴性菌落数各有一半(固体平皿法),这个生化特征趋向于剑川野鼠鼠疫的性质。结合表型与基因型的特征,本研究认为剑川50年代的菌株为野鼠鼠疫性质,据此,剑川野鼠疫源地的发现可以往前推至上个世纪50年代。
流行病学资料也显示,1950-1955年期间,剑川县累计发生人间鼠疫2次:1) 1950年,甸南镇浉河村下河,发病8人,死亡6人;2) 1954年,沙溪镇长乐村大长乐,发病34人,死亡14人。死亡率为47.6%,明显高出同一时期家鼠疫源地死亡率(21.1%)[1]。因此,还认为剑川野鼠鼠疫菌株对人的毒力强于家鼠鼠疫菌株。
3.3剑川野鼠鼠疫在云南鼠疫中的遗传定位 丽江鼠疫由青藏高原喜马拉雅旱獭鼠疫自然疫源地传入,而云南鼠疫起源于丽江鼠疫[15],并扩散至剑川形成野鼠鼠疫,进而再传至滇西及滇南广大区域而形成云南家鼠型鼠疫。本文中DFR+CRISPRs生成的MST也支持该观点。剑川野鼠鼠疫流行于上世纪50—90年代,虽然时间跨度数十年,但从MLVA结果来看,鼠疫菌种群变异度不大,形成了较稳定的克隆群,即便在流行中波及家鼠也未造成菌株遗传特征的改变。剑川野鼠簇中有2株菌分离自黄胸鼠(1954年长乐村,1984年石龙村),从基因组多态性位点来看,完全具备剑川野鼠菌株的基因特征,是野鼠鼠疫传至家鼠所致。
剑川菌株中,唯有YNJC20较为特殊,该菌1957年分离于甸南镇狮河村一褐家鼠体表寄生的绒鼠怪蚤,生化特征为麦芽糖发酵阳性,甘油发酵弱阳性[14]。在最初的DFR+CRISPRs分型中就以独立株的形式独立于各簇之外,与之最接近的不是剑川野鼠鼠疫簇而是家鼠鼠疫簇。将其分别与家鼠鼠疫簇和剑川野鼠簇进行VNTR位点比较,发现其与家鼠鼠疫簇的位点差异至少有10个,与剑川野鼠簇位点差异至少有14个,且该菌株天然缺失了pPCP质粒[16]。这体现了种群进化中的特殊变异。
综上,应该认为,云南剑川县境内发现2种类型的野鼠疫源地,除了以沙溪镇石龙为中心的原剑川野鼠疫源地,还有以金华镇庆华为疫点的另一新的野鼠疫源地。剑川野鼠疫源地的发现时间可往前推至上世纪50年代,同时疫情还扩散至家鼠和人群,因此长期认为剑川野鼠鼠疫从未波及人间的说法是不准确的。剑川野鼠鼠疫的长期存在,及2种类型疫源地的并存,增加了其防控的复杂性,因此应继续加强鼠间鼠疫的动态监测,及时发现和处置鼠间疫情,防止其波及到人间。
利益冲突:无
引用本文格式:石丽媛,丁奕博,张海鹏,等.云南省剑川县鼠疫疫源地分离菌株基因组多态性[J].中国人兽共患病学报,2020,36(1):20-24. DOI:10.3969/j.issn.1002-2694.2019.00.180
猜你喜欢
中国药学药品知识仓库(2021年12期)2021-12-07 00:43:43
东坡赤壁诗词(2019年5期)2019-11-14 10:36:10
十月(2019年4期)2019-07-29 17:33:30
云南档案(2019年5期)2019-06-20 09:16:36
中国学术期刊文摘(2019年23期)2019-01-29 11:49:47
绿色科技(2017年22期)2017-12-25 16:38:05
对联(2015年22期)2015-06-11 06:38:44
中国人兽共患病学报(2015年12期)2015-01-25 03:21:47
农业与技术(2014年3期)2014-04-25 09:13:07
农业与技术(2014年3期)2014-04-25 09:12:52
