
北京松山自然保护区中型捕食动物的食物构成分析
2020-03-12 01:06范雅倩张洪亮蒋万杰唐书培鲍伟东
生物学杂志 2020年1期
范雅倩,杨 婧,张洪亮,蒋 健,蒋万杰,唐书培,鲍伟东
(1. 北京松山国家级自然保护区管理处,北京 102115; 2. 北京林业大学 生物科学与技术学院,北京 100083)
分析野生动物食性可以了解动物的生存现状、食物组成季节变化、同类之间的竞争与共存关系、评价栖息地容纳量等诸多生态学内容[1-2],而且可以确定捕食者对不同猎物种类的选择压力,以及捕食效应对野生动物群落发展的影响,了解区域性野生动物多样性、食物链结构与功能的关系,进而提出针对动物种群和栖息地的有效保护管理措施。
北京松山国家级自然保护区是华北地区一处重要的野生动物富集区,入选原国家林业局划定的51处国家级自然保护区建设管理试点。自从晋升为国家级自然保护区后,得到了详细的本底调查[3-4],人们已对现有物种构成有了基本了解,统计有兽类14科29种,鸟类37科120种[5-6]。2010年以后使用红外相机对鸟类和兽类进行调查,又发现了以往未曾记录的亚洲狗獾(Melesleucurus)和松鼠(Sciurusvulgaris)的新分布[7-8]。这些本底资料为后续开展野生动物生态学研究、制定区域性生物多样性保护措施提供了坚实基础。
由于2022年冬奥会高山滑雪项目将在松山保护区附近举行,按照有关规定对保护区范围进行了重新调整,及时准确了解场地建设和栖息地转变对野生动物产生的干扰影响,是制定保护对策的重要内容。为此,本文基于以往调查积累,于2017年4月至11月利用动物粪样内容物鉴定法,对保护区3种食肉目动物进行了食物组成分析,以明确食物构成的差异,分析物种共存与竞争的生态学关系,为松山保护区野生动物管理提供基础资料。
1 研究区域与研究方法
1.1 研究区域
本研究在北京松山国家级自然保护区进行,该区拥有华北地区面积较大的天然油松林,是保存较为完好的暖温带山地森林生态系统,森林覆盖率87.7%,涵盖阔叶林、针阔混交林、针叶林[9],记载有脊椎动物68科216种及亚种[6]。2014年松山保护区经过调整后面积有所增加(6212 hm2),新增玉渡山区域面积2687 hm2,依然划分为2个核心区及其周边的缓冲区与实验区。本研究对亚洲狗獾(Melesleuculus)、貉(Nyctereutesprocyonoides)和豹猫(Prionailurusbengalensis)进行食物组成研究,在保护区全境进行动物遗留粪便样品的采集。
1.2 研究方法
北京松山保护区的森林植被生长良好,一些区域灌丛茂密,无法通行,因此,根据野生动物分布特点,以及能够开展外业工作的环境条件,选择山脊和林间能够行走的小道,设置动物粪样采集样线。在兰角沟与塘子沟监测区不同植被类型设置9条调查样线,在玉渡山范围内设置4条样线(图1)。
按照选定的样线,于春、秋季开展调查,以步行方式在天气晴好的条件下进行,收集调查样线上遇到的所有食肉动物粪便样品,记录样线上观察到的兽类动物足迹、啃食痕迹、毛发等活动信息,同时记录发现位点的GPS信息、测量样品的长度和宽度,如发现动物足迹则拍照记录。依据以往研究并参照文献确认动物种类[10-13],由于在本区域红外相机未拍摄到赤狐(Vulpesvulpes),犬科动物的粪样归为貉进行分析。貉的粪样以具针尾为特征;豹猫的粪便时常含有消化不完全的鸟羽而呈灰白色[14],两头圆钝形;亚洲狗獾的粪便内容物大多由昆虫外骨骼和植物果实组成,较粗、呈圆柱形[10]。当无法直接判断粪样归属时,则利用分子鉴定的途径予以准确区分[15]。
采用粪便内容物分析法对收集的样品进行分析,具体操作参照 Jedrzejewska 和 Jedrzejewski[16]的方法。依据猎物残余物的骨骼、毛发、牙齿等未消化成分尽可能鉴定到物种[17],并以获得上、下颌对数判断进食鼠类的个体数。以羽毛颜色、鸟喙和鸟爪形态与大小确定被食鸟类的物种以及数量(本研究仅划分为中型陆禽和小型鸣禽),依据剩余蝗虫和金龟子的残体计数昆虫残留物,进食植物不区分果实和茎叶。
2 结果与分析
本研究利用外部形态和DNA分子鉴定确认亚洲狗獾、貉、豹猫粪便样品 101 份,确认食物类型为 8 类,有食虫类(山东小麝鼩和川西缺齿鼩鼱)、野兔、松鼠类(岩松鼠、松鼠)、鼠类(黑线姬鼠、中华姬鼠、大林姬鼠、北社鼠、长尾仓鼠、棕色田鼠)、鸟类(勺鸡、鸫类、普通、鵐类)、宁波滑蜥、昆虫(蝗虫、金龟子、马陆)和植物。
食物种类的出现比率以小型哺乳类为主,鼠类的出现比例最高(56.7%),鸟类(12.7%)列在第 2 位,昆虫和植物列第 3、4 位。对野兔、勺鸡(上喙残体)、宁波滑蜥的捕食仅在亚洲狗獾各出现一例,在亚洲狗獾和貉样品中出现塑料垃圾和香烟过滤嘴。貉与豹猫的食物构成相似,但豹猫取食鸟类(25.7%)较多。
对食物成分的进一步分类发现,亚洲狗獾的食物中,林姬鼠(38.9%)和田鼠(16.7%)检出比率较高(表 1),尤其是对姬鼠的取食,在一份样品中检出 9 只个体的下门齿,推测是捡食其他猛禽取食剩余的头骨,原因是在该样品中,鼠毛很短,类似头部的毛发。由于小型鼠类个体小,尽管相对出现率较高(61.1%),但进食的生物量不高(占比29.1%),远不及野兔(47.2%)。在亚洲狗獾的食物中还检出蜥蜴鳞片和30条昆虫的幼虫,显示出亚洲狗獾的杂食性特点。貉与豹猫的食物构成相似,与这两种动物的运动能力一致较强,大型松鼠类动物提供的生物量占比较高,但豹猫对鸟类的捕食明显偏高。3种动物的食物中,昆虫和植物的重量贡献率均很低。
3种捕食动物的食物生态位宽度以豹猫最大、亚洲狗獾次之、貉较小为特征(表1),与豹猫捕食较多鸟类有关,而亚洲狗獾的捡食习性使其食物种类多于貉。食物谱重叠度指数表现出3种动物完全重叠(O=1.0),显示对共同食物资源的利用存在潜在竞争。
注:括号中数字为最多取食个体数
通过比较主要猎物相对重要值,小型鼠类、鸟类和野兔+松鼠构成了捕食者的常见食物(图 2)。除鼠类外,亚洲狗獾对其他食物的取食较为均衡。貉的食物中鸟类的重要值最低,无论出现率还是生物量均以小型种类为主,对貉的能量贡献率偏低,显示其运动能力不适合捕食鸟类。豹猫的食物中,除小型鼠类外,鸟类和野兔+松鼠类占有较高的重要值。
3 讨论
本研究中的3种动物包括了松山保护区中型食肉目动物的代表种类,因而它们的食性组成较为多样,植物叶占有较高比率,但是因食用的相对重量很低,在提供能量方面的作用较低,仅能够起到包裹消化道中猎物的毛发和碎骨,减少对消化道伤害的作用。尽管亚洲狗獾是食肉目动物,但昆虫也具有一定的相对重要值,与以往对北京市捕食动物的食物构成分析一致[11],但其所占比例偏低,不同于北欧狗獾(Melesmeles)的食物中无脊椎动物和蛙类的出现率较高的情况[23],显示出本区域的特点。本研究未采集猪獾(Arctonyxcollaris)的粪样,主要是由于其中的泥土较多,时常破解而无法采集,在发现地检视其中的成分,食物残渣以昆虫外壳和植物茎叶与种子居多,偶尔出现鸟类羽毛和鼠类牙齿,由此与亚洲狗獾出现食物资源分离。但由于昆虫资源提供的能量有限,不利于个体生存,这或许可以在一定程度上解释猪獾的红外相机拍摄率较高,即猪獾采取扩大取食范围的方式维持能量收入[8]。
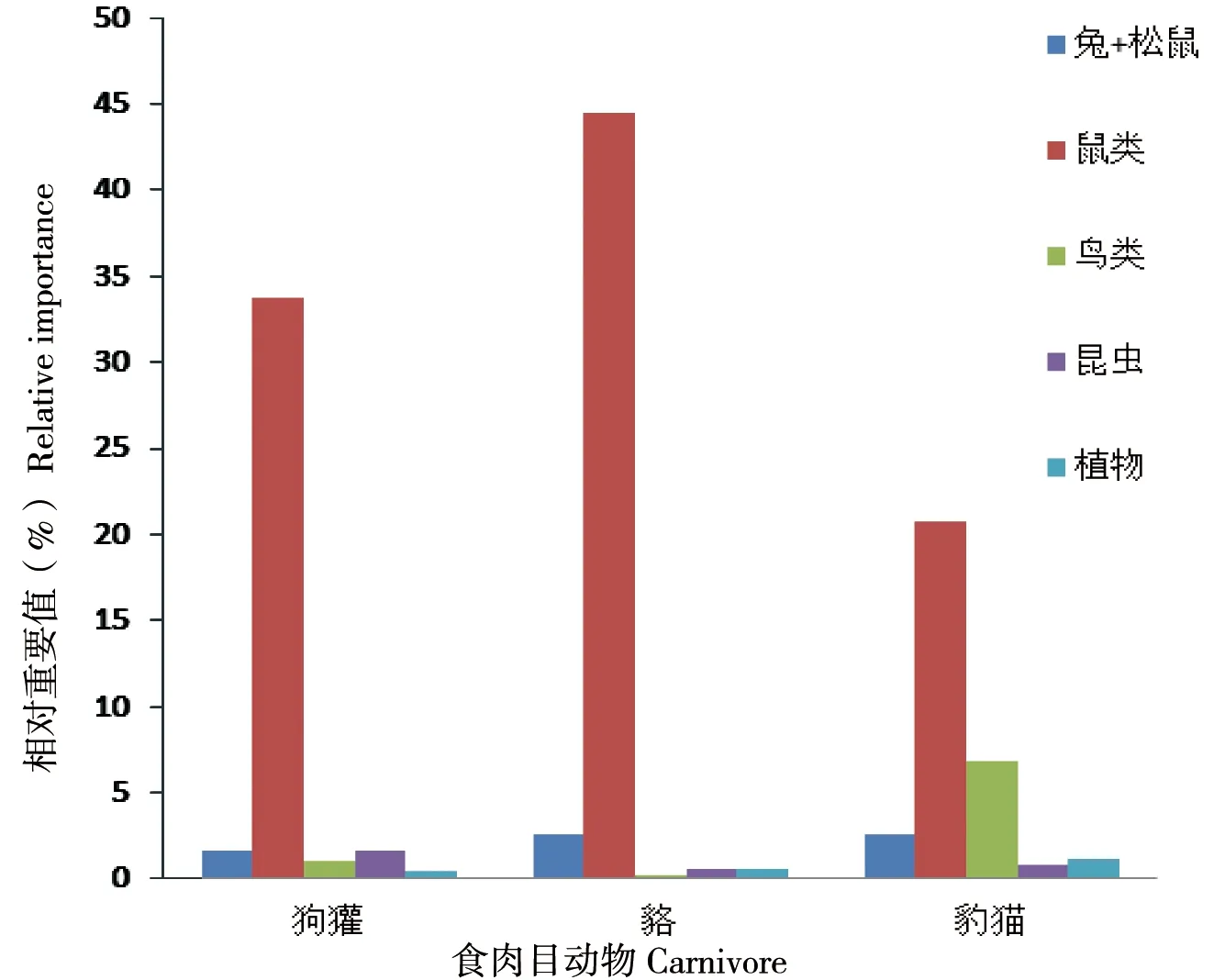
图2 北京松山3种食肉目动物主要食物相对重要值
在松山保护区豹猫的种群数量相对较多,收集到的粪样也最多,发现其食物构成具有一定特点,食物中鸟类的出现率高于亚洲狗獾和貉,鼠类也占有较高比例,但在重量比率上鼠类的贡献较大,而且松鼠类构成了鼠类猎物的主体,说明豹猫能够捕食运动灵活的松鼠,由此可以解释豹猫与其他动物在食物生态位上的差异,进而实现种间共存[24]。
3种捕食者的食物生态位宽度以豹猫最大、亚洲狗獾大于貉为特点,反映了运动能力和生物学特性影响食物组成的分离。对食物重叠度指数的计算表现出3种动物的重叠度极大,但从食物构成细分内容上看,豹猫与貉对同类食物的竞争最强,与两种动物主动捕食一致,这与来自其他区域的研究结论类似[25]。尽管3种捕食动物的食物生态位重叠度较高,但由于豹猫捕食较多鸟类,亚洲狗獾属于随机性捡食,貉取食小型啮齿动物,避免了种间直接竞争[23]。本研究发现的食物组成特点,为后续深入开展捕食动物种间共存提供了基础信息,将有助于提高松山自然保护区食肉类动物的生态管理。
猜你喜欢
林业科技(2021年2期)2021-07-08
野生动物学报(2018年4期)2018-11-23
小天使·一年级语数英综合(2018年5期)2018-06-22
小天使·一年级语数英综合(2017年9期)2017-10-20
小天使·一年级语数英综合(2017年2期)2017-02-16
小天使·一年级语数英综合(2017年2期)2017-02-16
中学理科园地(2016年6期)2017-01-03
科技视界(2016年23期)2016-11-04
现代农业科技(2009年11期)2009-07-08
