
基于光合电子流的超级早稻光合特性研究
2020-03-11 08:34:12贺晓鹏边建民朱昌兰傅军如贺浩华
核农学报 2020年2期
贺 俐 罗 辉 贺晓鹏 边建民 朱昌兰傅军如 吴 杨 贺浩华
(1江西农业大学作物生理生态与遗传育种教育部重点实验室/江西省作物生理生态与遗传育种重点实验室/农学院,江西 南昌 330045;2井冈山大学生命科学学院,江西 吉安 343009)
水稻(Oryza sativaL.)作为一种重要的粮食作物,为了满足人类对粮食的需求,其单产的提高刻不容缓,当前超级稻培育是提高产量的重要途径[1-2]。光合作用是作物生产力高低的关键因素[3],因此,越来越多科研工作者关注水稻的光合特性研究,主要集中在不同生态因子对水稻光合特性影响[4-7],不同栽培方式对水稻光合特性的影响[8-9],比较不同水稻品种间的光合特性[10-11]等方面。研究中常见的光合特性生理指标有饱和光强(saturation light intensity of photosynthesis rate,Ⅰsat)、净光合速率(net photosynthetic rate,Pn)、PSⅡ有效光量子效率(actual photochemical efficiency in photosystem Ⅱ,ΦPSⅡ)等,很少有关于光合电子传递速率(electron transport rate,ETR)的研究报道。与光合电子流有关的研究中,一般用光系统Ⅱ的ETR 来代表光合电子流的大小,其值越大,表明其形成光合能力越强[12]。近些年,光合ETR与光强之间的关系及其在植物光合过程中的作用得到了科研人员的广泛关注,并取得了较大进展[13-15]。
在超级稻光合特性研究中,大量试验都是比较不同光合指标在品种间的差异或是不同环境条件下指标变化的趋势来研究品种间的光合特性[16]。而植物光合作用原初反应过程中电子传递相关的参数,最大电子传递速率(ETRmax)、饱和光强(Saturation light intensity of electron transport rate,PARsat)、初始斜率(αe)等一般通过光合模型拟合计算[17]。不同模型拟合效果相差较大,因此参数的拟合计算非常关键,模型的选择直接影响后续试验的开展。目前,植物光合电子流对光响应模型主要分为经验模型和半机理模型两大类[14]。其中,经验模型的建立没有考虑到植物光合作用的原初反应、电子传递等过程,且不能合理解释光抑制现象,无法直接计算光饱和点[12]。半机理模型只涉及光能的吸收和激发,未考虑电子的共振传递和转化等过程,同样不涉及捕光色素分子内禀参数在这些过程中的作用,无法计算ETRmax和PARsat等光合参数[18-19]。鉴于此,在充分考虑光合作用的原初反应过程和捕光色素分子光能吸收特性的基础上,Ye等[20-21]建立了一个光合电子流对光响应的机理模型。该模型已得到国内外学者的广泛认可和引用[22-23]。
植物叶片捕光色素分子对光能的吸收、传递以及电荷分离等过程与光合作用的原初反应密切相关[24-25]。在正常环境下植物的光合作用是由植物本身的内禀特性所决定的[26],如捕光色素分子的数量、光能吸收截面和光能退激发速率等因素[27]。原初反应过程中形成的光合电子流是植物光合能力形成的重要基础,然而植物叶片中的哪些内禀特性会对ETR 产生影响并进一步影响其光合能力鲜有报道。因此,本研究从光合电子流对光响应的机理模型出发[20-21],测定并推算光合电子流相关特征参数及捕光色素物理参数,研究相同环境条件下超级早稻的光合特性,探索超级早稻叶片的ETR 对光响应的变化规律,揭示光合电子流与光强、超级稻捕光色素分子物理特性之间的关系,旨在为超级早稻的光合作用机理研究提供重要的理论依据。
1 材料与方法
1.1 试验材料与试验地概况
本研究于2016 年3-7 月在江西省吉安市青原区雷家村(27°11′N,115°02′E)成片的双季稻水稻生产田内进行。年平均气温17.7℃,全年≥10.0℃活动积温为5 500 ~5 800℃;全年≥10.0℃积温天数250 ~300 d;年平均降雨量1 487 mm;年日照时数平均为1 814 h。土壤理化性质为:pH 值5.61、有机质2.49 g·kg-1、全氮1.07 g·kg-1、碱解氮141.28 mg·kg-1、速效钾67.12 mg·kg-1、有效磷14.86 mg·kg-1。供试材料分别为淦鑫203、金优458、株两优819、春光1 号、荣优9 号、荣优286、03 优66;对照:金优402,均由江西省作物生理生态与遗传育种重点实验室提供。
1.2 试验设计
试验早稻在土壤肥力相同的同一试验田栽培,采用随机区组设计,小区面积为16 m2,3 次重复。大田育秧,3 月24 日播种,4 月24 日移栽,行株距为26.4 cm×15 cm,基本苗2 株/穴。施纯N 180 kg·hm-2,P2O5105 kg·hm-2,K2O 180 kg·hm-2。氮肥施用比例为基肥∶分孽肥∶穗肥=4∶3∶3;磷肥全部作为基肥;钾肥施用比例为基肥∶分孽肥∶穗肥=4∶3∶3。基肥在移栽前2 d 使用,在移栽后5~7 d 施用分孽肥,在倒三叶露尖时施用穗肥,其他大田管理措施按照高产农田要求进行。于6 月26 日起连续3 d 晴天进行光响应曲线的测定,早稻处于灌浆期,随机选取3 株长势相当的健康植株,每株选择3 张剑叶进行测量。
1.3 测定项目与方法
1.3.1 光合电子流对光响应数据的测定 在晴天上午8:00-12:00,利用LI-6400-40B 便携式光合仪(Li-Cor INC,美国)自带光源,设自动测量程序进行数据测量,利用CO2钢瓶控制CO2泵流速为500 mmol·s-1,浓度为390±2 μmol·mol-1,相对湿度控制在50%~70%,叶室温度设定为35±1℃。光合有效辐射梯度设置为:2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50 和0 μmol·m-2·s-1,每隔180 s 记录一个点,测定叶片在不同光强下ETR 等光合参数。利用Ye 等[20-21]的机理模型对光合电子流响应曲线进行拟合,计算ETRmax、PARsat、αe等。
1.3.2 捕光色素分子内禀参数的计算 捕光色素分子的有效光能吸收截面(effetive light absorption crosssection,σ′ik)、本征光能吸收截面(eign-absorption cross-section,σik)、捕光色素分子数(number of lightcatching pigment molecules,No)处于最低激发态的捕光色素分子数(light-harvesting pigment molecules,Nk)、捕光色素分子处于最低激发态的最小平均寿命(the shorter minimum overage lifetime,τmin)的计算参照Ye 等[20-21]的方法。
1.3.3 叶绿素含量的测定 参照Arnon[28]的方法并加以改进。用打孔器(直径5 mm)避开叶脉打15 个圆片,剪碎后置于离心管内,加入5 mL 80%丙酮,在黑暗处浸提24 h,用UV-2550 紫外可见分光光度计(日本岛津)分别测定470、645、652、663 nm 波长处的吸光度值,并根据吸光度值计算单位面积的叶绿素含量。
1.4 数据分析
采用“光合计算软件”(http:/ /photosynthetic.sinaapp.com/calc.html)对测量数据进行分析。利用SPSS 22.0 软件对试验数据进行统计学分析,OriginPro 8.5 软件作图。
2 结果与分析
2.1 光合电子传递速率(ETR)对光强的响应曲线
由图1 可知,Ye 等[20-21]建立的光合电子流对光响应的机理模型很好地拟合了不同光强下水稻叶片的ETR-Ⅰ曲线,所有早稻品种的叶片在光强低于400 μmol·m-2·s-1时,ETR 随光强的增加几乎直线增加,超级早稻与CK 的ETR 无显著差异(P<0.05);在光照强度400~1 600 μmol·m-2·s-1范围内,随着光强的增强,所有水稻品种的ETR 快速升高,超级早稻的ETR显著高于CK(P<0.05)。当升至设置的最大光强(2 000 μmol·m-2·s-1)时,所有水稻品种的ETR 均表现出下降趋势,出现PSⅡ的动力学下调现象[29]。与CK 相比,超级早稻保持着较高的ETR,具有更强的光合能力。
由表1 可知,ETRmax和PARsat的拟合值与实测值非常接近,R2超过0.999。CK 的初始斜率αe最大,为0.351 μmol·μmol-1,显著高于其他超级早稻品种,说明其把光能转化为电子流的能力最强,但所有超级早稻品种的ETRmax均显著高于CK,其中春光1 号的ETRmax最大,为228.45 μmol·m-2·s-1。CK 的PARsat最小,株两优819 的PARsat与CK 无显著差异,其他超级早稻品种的PARsat均显著高于CK,淦鑫203 的PARsat最大,为1 659.12 μmol·m-2·s-1。

图1 超级早稻叶片的ETR 对光强的响应曲线Fig.1 Light response curves of electron transport rate for leaves of super early rice

表1 超级早稻光合参数的测量值与拟合值Table 1 Measured and fitted values of photosynthetic parameters for super early rice
2.2 光合叶绿素含量及捕光色素分子内禀特性的比较
由表2 可知,不同早稻品种间的叶绿素含量存在着差异,其中CK 的叶绿素含量最高,为702.36 mg·m-2,淦鑫203 的叶绿素含量最低,为558.07 mg·m-2,除春光1 号外,其他超级早稻的叶绿素含量均显著低于CK。除了ETRmax和PARsat等光合参数,通过光合电子流对光响应机理模型还可估算捕光色素分子的一些物理参数,如N0、τmin、σik等,超级早稻叶片中N0的变化情况和叶绿素相似,除超级早稻春光1 号外,其他超级早稻的No均显著低于CK。CK 的叶绿素含量最高,但其捕光色素分子的σik最小,为1.81×10-21m2,淦鑫203 的叶绿素含量最低,但其捕光色素分子的σik最大,为2.32×10-21m2,春光1 号的σik与CK 相比无显著差异,其他超级早稻品种的σik显著高于CK。CK 的τmin最长,为132.95 ms,显著高于其他超级早稻品种。

表2 超级早稻叶片的叶绿素含量和捕光色素分子物理参数Table 2 Chlorophyll content and physical parameters of light-harvesting pigment molecules for super early rice
2.3 不同光照强度下的捕光色素分子有效光能吸收截面
光能吸收截面能反映植物捕光色素分子对光能吸收能力的强弱,捕光色素分子的σ′ik则表示其在不同光强下捕获光子能力的大小[20-21]。由图2 可知,各早稻品种的σ′ik随光强的增加呈非线性下降,但各早稻品种σ′ik的下降速度有所不同,其中淦鑫203 和株两优819 的σ′ik下降速度较慢且非常接近,CK 的σ′ik下降最快,σ′ik减少得越慢,越有利于光能的吸收和利用,上述结果表明,与CK 相比,超级早稻在不同光强下仍然保持着较大的σ′ik,特别是在高光强区域,淦鑫203的σ′ik最大,超级早稻品种的σ′ik均高于CK,说明超级早稻品种在强光下仍然保持着较强的光吸收能力。
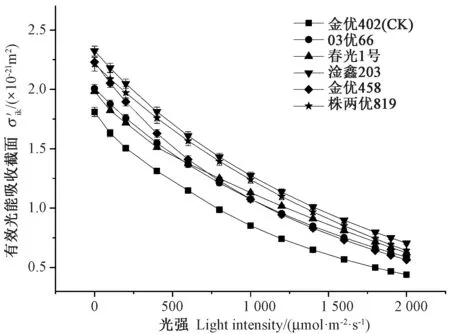
图2 超级早稻的有效光能吸收截面对光强的响应曲线Fig.2 Light response of curves of theeffective light absorption cross-section versus PAR for super early rice
2.4 处于最低激发态的捕光色素分子数对光强的响应曲线
随光强的不断增加,所有早稻品种的Nk均呈非线性增加的趋势,一旦处于激发态的捕光色素分子不能及时回到基态,这些激发态的捕光色素分子将对植物的光合器官产生伤害,从而影响植物光能的吸收和利用[30-31]。由图3 可知,随着光强增加,CK 的Nk上升最快,淦鑫203 的Nk上升最慢。在不同光强下,各超级早稻品种与CK 相比均保持着较低水平的Nk,表明超级早稻品种中有较多的捕光色素分子处于基态,有利于其吸收光能。在高光强处,超级早稻的Nk均小于CK,表明超级早稻在高光强时的光吸收能力较CK 强。

图3 超级早稻的有处于最低激发态的捕光色素分子对光强的响应曲线Fig.3 Light response of curves of photosyntheticpigment numbers in the lowest excited state versus PAR for super early rice
3 讨论
植物的光合能力与原初反应过程中光合电子流的形成密不可分,光合电子流不仅是光合能力形成的基础,还是光合碳同化过程中的能量源泉[14]。原初反应过程中捕光色素分子的内禀特性不仅决定了光能的吸收与传递,也将影响到其后激发能的分配[32]。植物叶片的光合特性与捕光色素分子光能的吸收、传递和电荷分离等过程密切相关[27]。本研究运用光合电子流对光响应的机理模型从ETR 和捕光色素分子内禀特性两个角度对超级早稻叶片的光合特性差异进行了探讨。结果表明,不同光强下盒超级早稻叶片ETR 对光的响应曲线相类似,在低光强处,随着光强的增加超级早稻叶片ETR 快速增加,ETR 达到最大值后随着光强的增加而缓慢下降,存在非常明显的PSⅡ动力学下调[29],这与捕光色素分子的σ′ik随光强增加而下降有关[12]。本研究运用的光合电子流对光响应的机理模型很好地拟合了超级早稻叶片的光合电子流对光的响应曲线,具有极值,存在拐点,可以准确地得到ETRmax和PARsat。本研究结果显示ETRmax和PARsat的拟合值与实测值非常接近,这与前人研究结论一致[33-35]。
通过测定和推算与光合作用相关的一些参数,可以反映植物对光的捕获和利用能力。植物的ETR 与1,5-二磷酸核酮糖(Ribulose bisphosphate,RuBp)的再生能力密切相关,较低的ETR 会限制植物体内NADPH 的生成与光合磷酸化,进而限制植物的最大羧化效率(maximum carboxylation rate,Vcmax)[36],最终降低其光合能力。王强等[37]对濒危植物长叶榧进行光合生理生态研究发现,夏季长叶榧较强的光合作用与其具有较高的ETRmax有关。本研究结果表示,超级早稻保持着较高的ETR,其ETRmax显著高于CK,具有更强的光合能力。PARsat反映了植物对不同光强的利用能力和适应性。叶子飘等[38]对不同草本植物光合特性差异进行研究,发现大狼把草的PARsat显著高于山莴苣,且具有更高的ETRmax,其光适应能力更强。本研究发现,超级早稻品种的PARsat显著高于CK,表明超级早稻具有更强的光合适应能力。
植物叶片的叶绿素含量与其光合能力密切相关。植物叶片的ETRmax和PARsat不仅与叶绿素含量有关,还与捕光色素分子的内禀特性有关[26,38]。本研究中,超级早稻的叶绿素含量均低于CK,但其ETRmax和PARsat均高于CK,这可能与超级早稻具有较大的σik和较短的τmin有关。σik能反映植物捕光色素分子的潜在光能吸收能力,σik越大,捕光色素分子吸收光子的能力越强,植物吸收光能的能力越强[32]。τmin值越小,激子越容易退激发到稳定的基态,越有利于能量传递到光化学反应中心,ETR 就越大[17]。因此,单纯地认为叶绿素含量越多,ETRmax和PARsat就越大,植物光能吸收和利用能力就越强具有片面性,还要考虑植物捕光色素分子的σik、τmin等内禀参数的影响。
植物的光能吸收截面与其所处环境密切相关[19]。捕光色素分子的σ′ik表示捕光色素分子在不同光强下的实际光能吸收能力[21]。σ′ik随光强的增加而下降,不同植物的σ′ik对光的响应是不同[26,39]。本研究中,超级早稻的捕光色素分子的σ′ik随光强的增加下降速度较缓慢,而CK 的捕光色素分子的σ′ik随光强的增加下降速度较快,表明超级早稻的捕光色素分子在光强较弱时的光能吸收能力较强,而CK 的捕光色素分子光能吸收能力较弱,在高光强区域,超级早稻可以吸收更多的光子使光系统Ⅱ产生电荷分离,其ETR 高于CK。捕光色素分子吸收光能后,从基态跃迁到激发态。处于高能态的捕光色素分子将通过共振传递的方式将大部分能量传递到光反应中心,使PSⅡ发生电离,产生光合电子流,剩余的能量则通过荧光退激发和热耗散消耗掉[27,40-41]。三种退激发方式间存在着竞争,如果过多的捕光色素分子处于激发态且不能及时回到基态,过多的捕光色素分子且处于激发态容易产生大量活性氧,与水作用形成H2O2,最终破坏光合结构,且处于基态的捕光色素分子数骤减,植物的光能吸收和电子传递受到阻碍,严重影响光合电子流的产生和后续的碳同化作用[40,42],从而对植物产生伤害。本研究结果表明,在不同光强下,各超级早稻品种的Nk均低于CK,表明超级早稻品种具有较多的捕光色素分子处于基态,有利于其光合电子流的产生和光合作用的进行。这可能与超级早稻的τmin均小于CK 有关,τmin越小,处于基态的捕光色素分子越多,越有利于光能的吸收和利用。
4 结论
本研究以超级早稻为试验材料,通过测定不同光强下超级稻的ETR 等光合参数,首次利用光合电子流对光响应模型拟合了超级早稻的光合电子流光响应的曲线并对品种间光合特性参数的差异进行了分析。结果表明,超级早稻相比CK 具有较好的光合特性,表现出ETRmax大,PARsat高的特性。水稻对光的捕获和利用能力是影响其光合作用的一个决定性因素,水稻捕光色素分子的内禀特性与光合特性密切相关。此外,捕光色素分子的σ′ik和处于激发态的捕光色素分子对光响应曲线的差异也有可能影响它们的光合特性。本研究仅从光合电子流的角度对超级稻的光合特性进行了分析,今后还可以从净光合速率对光响应、光能利用效率、光化学量子效率等角度对其进行全面的分析。
猜你喜欢
农产品市场周刊(2022年14期)2022-08-10 07:09:10
农业科技与信息(2021年23期)2021-12-06 01:14:10
今日农业(2021年17期)2021-11-26 23:38:44
古今农业(2021年1期)2021-07-20 07:52:08
制造技术与机床(2019年6期)2019-06-25 10:18:08
中国新技术新产品(2019年21期)2019-01-20 02:41:59
电子技术与软件工程(2017年23期)2018-01-17 08:10:50
科学家(2016年12期)2017-09-29 02:15:14
中国市场(2017年13期)2017-06-03 23:57:00
现代制造技术与装备(2015年4期)2015-12-23 10:20:20
