
三亚大东海砂质潮间带小型底栖生物的时空分布及影响因素
2020-03-08 14:00梁宸梓杨丽莉李亚男慕芳红
海洋科学 2020年2期
骆 添, 梁宸梓, 孙 燕, 杨丽莉, 李亚男, 臧 瑜, 慕芳红
三亚大东海砂质潮间带小型底栖生物的时空分布及影响因素
骆 添, 梁宸梓, 孙 燕, 杨丽莉, 李亚男, 臧 瑜, 慕芳红
(中国海洋大学 海洋生命学院, 山东 青岛 266003)
于2015年和2016年对三亚大东海砂质潮间带进行了季节性的调查, 对小型底栖生物及其所处的沉积环境进行了研究, 探讨了小型底栖生物的丰度、生物量和群落的时空变化及其与环境变量的关系。共鉴定小型底栖生物类群11个, 优势类群为海洋线虫类与桡足类(二者占总丰度83.13%); 小型底栖生物年平均丰度为(595.77±442.72)ind·10cm–2, 小型底栖生物2015年12月的平均丰度为(768.70± 533.22) ind·10cm–2, 2016年6月平均丰度为(422.85±236.42) ind·10cm–2。丰度分布呈现明显的季节和潮区差异。相关分析结果表明pH、偏态系数与小型底栖生物的丰度呈极显著的负相关, 分选系数、中值粒径与之呈极显著的正相关性。小型底栖生物的丰度随季节变化趋势为冬季>夏季。食物丰度、溶氧量升高与沉积物粒径的变化是冬季桡足类丰度的大幅增加的主要原因。TWO-WAY ANOVA与ANOSIM结果显示, 潮带对小型底栖生物丰度和群落结构的影响在冬季显著。SIMPER分析结果显示, 此差异主要来源为桡足类和线虫类。BIOENV分析结果显示, 溶氧量和分选系数的组合最能解释这种差异。这一结果可能是由于高、中潮带的人为干扰与砂滩的清理作用导致。夏季潮带间丰度与群落结构差异不显著, pH、盐度、叶绿素含量是影响其分布的主要原因。研究结果可为砂质滩小型底栖生物的研究提供基础数据, 也可为全球气候变化背景下砂质潮间带的保护、开发和利用提供科学依据。
小型底栖生物; 时空分布; 丰度; 群落结构; 大东海; 三亚
小型底栖生物是指是可被0.031~0.5 mm筛网截留的底栖生物类群, 是海洋碎屑食物链的重要环节, 是仔、稚、幼鱼和虾蟹等经济动物的重要饵料; 此外, 由于其具有分布广泛、物种丰富度高、繁殖力强、生命周期短、对自然和人为的影响的响应明显等特点, 可作为环境监测的生物类群。作为陆地和海洋的分界面之一, 砂质潮间带具重要的生态功能和生态服务价值, 但却长期被忽视。
砂质潮间带是典型的由砂和波浪能等物理因素驱动的生态系统[1]。栖息于砂滩的生物大多营隐蔽的挖掘或者间隙生活, 不为人所见, 砂质滩一度被认为是“生态沙漠”, 但实际上它拥有丰富多彩的生物群落, 栖息着极为多样的底栖动物[2]。当前砂质潮间带面临着人类扰动和全球变化的双重威胁, 特别是海平面上升和海岸侵蚀、海洋表层的暖化和酸化, 将在大尺度上深刻地改变其生态系统和生物多样性[3]。
三亚市海岸线长209.1 km, 沿岸滩涂25.1 km[4], 然而关于该地区, 特别是砂质潮间带小型底栖生物的相关报道却为空白。本研究选取三亚大东海砂质潮间带为野外采样地点, 对小型底栖生物的丰度、类群与群落结构三个方面, 进行分布趋势及其与环境因子间关系两个层次的分析, 研究结果可为三亚大东海潮间带地区生物资源的可持续利用提供基本依据, 同时为全球气候变化背景下砂质潮间带的保护、开发和利用提供科学依据。
1 材料与方法
1.1 取样地点与方法
本研究用PVC 取样管(D=4.4 cm)在三亚大东海砂质潮间带(18°13′19″N, 109°31′0″E)进行冬季(2015年12月)与夏季(2016年6月)小型底栖生物以及环境因子的采集和原位测定。
断面与潮带的组合即为一个采样点(如断面1高潮带), 每个采样点采取三管20 cm深的沉积物芯样用于小型底栖生物的分选, 芯样采出后立即装入500 mL样品瓶中, 并加入等体积的10%甲醛溶液, 混匀密封后常温保存。每个采样点另采两管20 cm的沉积物芯样装入封口袋中, 用于粒度、有机质、叶绿素等的测定, –20℃冷冻保存。

图1 采样地点(审图号: 9122435998)
1.2 样品处理方法
生物样品: 虎红染色, 0.031 mm 和 0.5 mm孔径的网筛过滤, 按小型底栖生物类群分别挑选计数[5, 6]。
环境因子测定样品: 采用筛析法测定粒度, 并计算中值粒径(MDΦ)、分选系数(QDΦ)、偏态系数(SKΦ)。根据《海洋调查规范》[7]采用荧光分光光度法测定并计算叶绿素a (Chl)的含量[8], 通过重铬酸钾氧化-还原容量法测定有机碳含量。
原位测定: 使用多参数水质仪(In-Situ Inc, Smartroll MP)测定每个采样点沉积物间隙水的水温、pH, 盐度、溶解氧含量。
1.3 数据处理与分析
生物量的估算采用类群的平均个体干重经验值乘以各类群丰度的方法[9-10], 各类群分别采用的个体平均干重见表1[6, 11-12]。采用SPSS对数据进行统计分析。以百分比含量表示的环境因子数据经对数转换, 采用R语言对环境因子进行PCA分析, 得到样本之间的主要差异来源。

表1 小型底栖生物的个体平均干重[9-10]
使用SPSS进行ONE-WAY ANOVA和TWO-WAY ANOVA分析, 对丰度和生物量的时空分布进行差异显著性检验, 之后对丰度和环境因子作相关性分析。群落结构分析使用R语言ade4和vegan软件包, 在样本数据基础上建立Bray-Curtis距离矩阵, 对潮区和季节两个因素进行ANOSIM检验。之后进行SIMPER分析得到各类群对分组的贡献, 并对群落数据和环境因子数据进行BIOENV分析, 得到最能解释丰度组成的空间异质性的环境因子组合。
2 结果与分析
2.1 沉积环境
对本研究所调查的三亚大东海潮间带环境因子进行测量和分析, 结果详见表2。PCA分析结果如图2所示。PC1与PC2 的累计信息量为67.6%, 可以较好的对环境因子间的关系进行描述。对PC1贡献较大的依次为pH, 温度, 偏态系数, 溶氧量, 盐度, 分选系数, 有机质, 叶绿素含量, 中值粒径; 对PC2贡献较大的依次为叶绿素含量, 偏态系数, 分选系数, pH, 温度, 中值粒径, 有机质, 盐度, 溶氧量。
2.2 小型底栖生物类群组成
2个季节采样的小型底栖生物共鉴定出11个类群, 包括多毛类、腹毛类、寡毛类、介形类、昆虫类、螨类、桡足类、线虫、涡虫、无节幼虫类以及其他未鉴定类群。
小型底栖生物的平均丰度为(595.77± 442.72)ind·10cm–2。小型底栖生物各主要类群的平均丰度、生物量见表3。

表2 采样点环境因子

图2 采样点环境因子PCA分析
注: 高: 高潮带; 中: 中潮带; 低: 低潮带; 1: 1断面; 2: 2断面
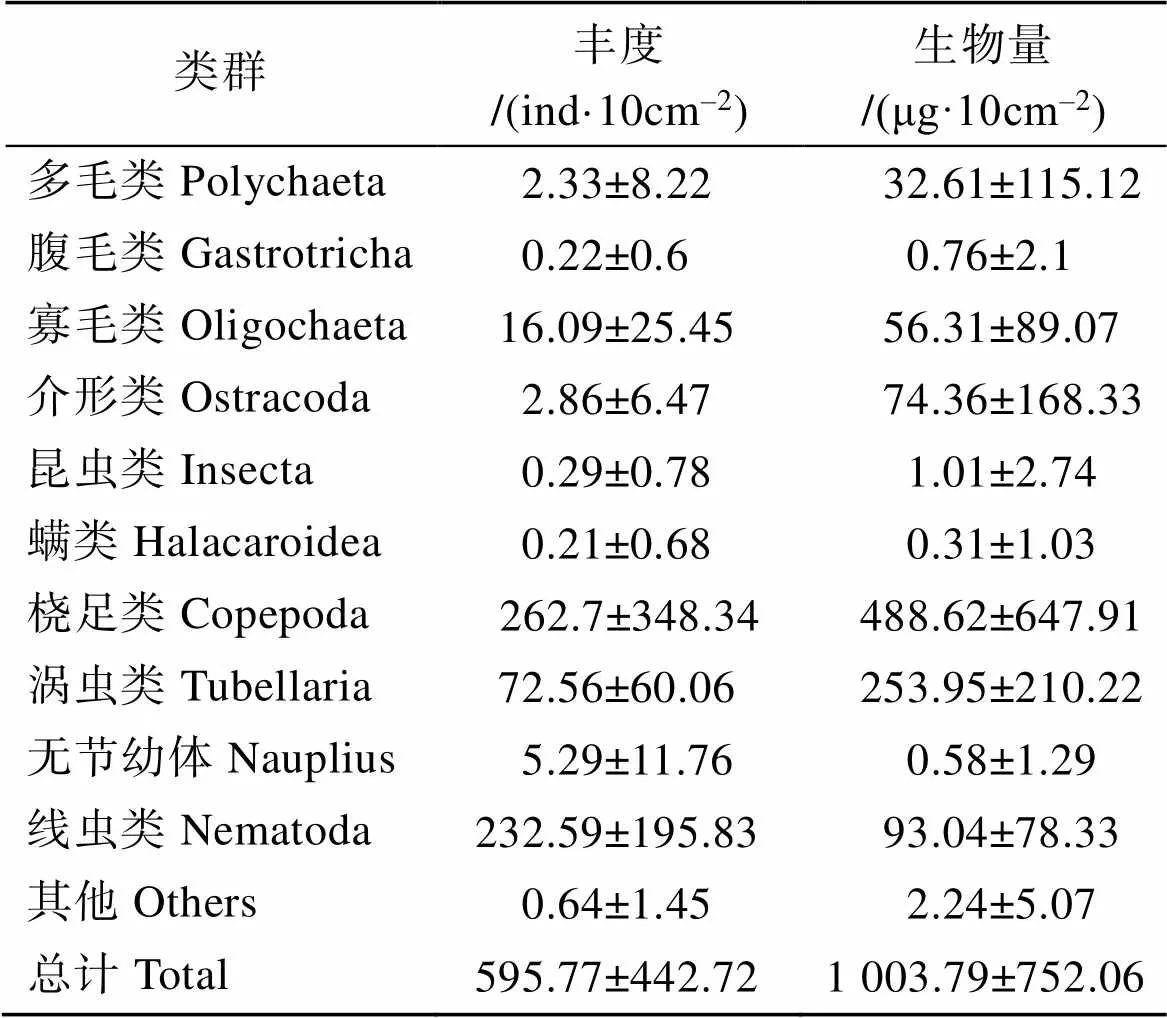
表3 小型底栖生物各主要类群的平均丰度、生物量
小型底栖生物丰度百分比组成如图3a所示, 桡足类优势度最高, 线虫居次。其他依次为涡虫、寡毛、无节幼体类。另外6个类群, 包括介形类、多毛类、昆虫类、腹毛类、螨类和其他未鉴定类群合计约占总丰度的1.10%。

图3 小型底栖生物主要类群
注: a: 丰度; b: 生物量
生物量百分比组成如图3b所示, 桡足类最多, 涡虫类居次, 其他依次为线虫、介形类、寡毛类。其他6个类群合计约占总生物量的3.74%。
2.3 小型底栖生物丰度与生物量分布及其与环境因子的关系
2.3.1 丰度与生物量的时空分布
夏季小型底栖生物平均丰度为(422.85±236.42) ind·10cm–2, 最高值出现在高潮带(图4a)。在高、中潮带桡足类是优势类群。夏季平均生物量为(903.90± 651.42) μg(dwt)·10cm–2, 高潮带>中潮带>低潮带(图4b)。
冬季小型底栖生物的平均丰度为(768.70± 533.22) ind·10cm–2, 最高值出现在低潮带(图4c)。线虫在高、中潮带是优势类群, 分布趋势不随潮带变化; 桡足类丰度低潮带最高; 其他类群的丰度无潮区差异。冬夏季小型底栖生物总丰度的差异主要由于冬季低潮带桡足类丰度的大幅增加。冬季平均生物量为(1103.68±847.92) μg(dwt)·10cm–2, 低潮带生物量最高(图4d)。
对各站位丰度数据用SPSS进行正态性与方差齐次性检验, 方差不齐。对数据进行开方转换后, 分别对潮带和季节进行ONE-WAY ANOVA分析, 并对此二因素进行TWO-WAY ANOVA分析, 结果(表4)显示, 在高潮带和中潮带, 季节对丰度的影响不显著, 但是在低潮带季节的影响显著(<0.05); 冬季, 潮带对丰度的影响显著(< 0.05)。由成对比较结果可以看出, 冬季的高潮带与中潮带之间并不存在显著差异, 但二者与低潮带之间具有显著差异(<0.05)。而夏季, 潮带之间的小型底栖生物丰度并未表现出显著差异。
表4结果显示, 在高潮带和低潮带, 季节对生物量的影响显著(<0.05), 在中潮带季节性差异不显著。冬季, 潮带对生物量的影响显著(< 0.05)。由成对比较结果可以看出, 冬季的高潮带与中潮带之间并不存在显著差异, 但二者与低潮带之间具有显著差异(<0.05)。夏季, 高潮带与低潮带之间有显著差异。
2.3.2 丰度与环境因子相关性
用R语言对丰度和所有环境因子进行正态检验, 结果除了溶氧量、叶绿素含量和 QD外其他值都不符合正态假设。对数据进行非参数Spearman相关分析。

表4 基于潮带与季节的丰度与生物量的时空分布差异(P值)
注:<0.05: 差异显著
由表5可以看出pH、偏态系数与小型底栖生物的丰度呈极显著的负相关, 分选系数、中值粒径与其呈极显著的正相关。温度、pH、偏态系数与线虫的丰度呈极显著的负相关, 盐度、有机质含量、叶绿素、分选系数、中值粒径与其呈极显著的正相关。盐度与桡足类丰度呈极显著的负相关, 分选系数、中值粒径与其呈极显著的正相关。其他环境因子与三者的相关性不显著。
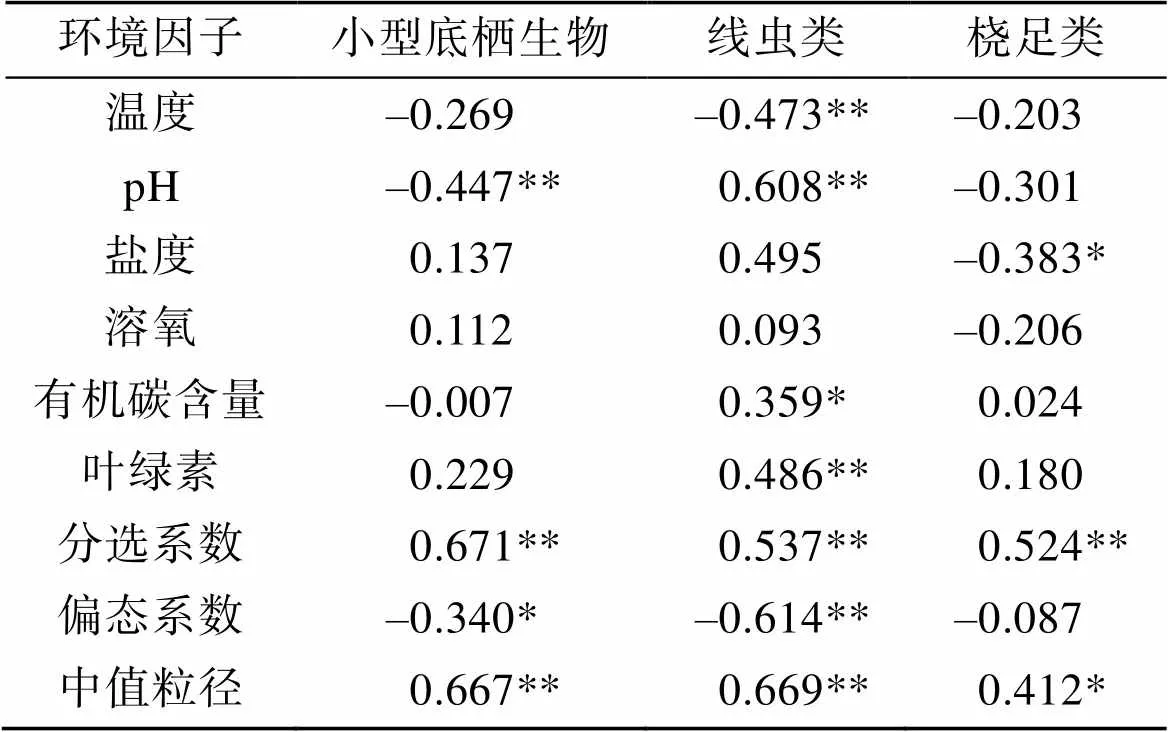
表5 小型底栖生物丰度与环境因子的相关关系
注: *:<0.05; **:<0.01
2.4 群落空间异质性及其与环境因子的关系
ANOSIM分析结果显示小型底栖生物群落在季节之间差异显著(=0.1806,0.005<0.05)。分别对冬夏两季的小型底栖生物类群丰度进行ANOSIM分析, 结果显示冬季潮带间差异显著(=0.630 9,= 0.001<0.05), 夏季潮带间差异不显著。

图5 各类群对冬季潮带间群落空间异质性的贡献
注: a: 中潮带与低潮带; b: 高潮带与低潮带; c: 高潮带与中潮带
SIMPER分析结果显示造成冬季潮带间群落异质性的主要类群是桡足类和线虫类, 贡献低潮带与中高潮带间50%以上的差异。BIOENV分析结果显示, 溶氧量和分选系数的组合最能解释冬季潮间带小型底栖生物群落结构, 其相关系数为0.4260。叶绿素含量和有机质的组合最能解释夏季潮间带小型底栖生物群落结构, 其相关系数为0.3455, pH、盐度、叶绿素含量最能解释低潮带小型底栖生物群落结构随季节的变化, 其相关系数为0.5562。
3 讨论
小型底栖动物丰度及波动受到很多因素的影响, 水温、水深、季节、叶绿素含量、溶氧量、沉积物粒度、有机质含量、以及自身繁殖特点等[13-17]。华尔[18]等人的研究表明, 砂质潮间带小型底栖生物随纬度的大尺度分布格局主要受到温度带梯度, 沉积物粒径, 盐度以及溶氧量的影响。与中国海砂质潮间带小型底栖生物历史研究数据比较[19-27], 三亚大东海小型底栖生物丰度相对较低, 与华尔等[18]研究得出的亚热带潮间带小型底栖生物丰度通常低于温带的规律一致。
3.1 丰度与生物量的季节变化及其与环境因子的关系
本研究小型底栖生物的丰度存在显著的季节变化, 表现为冬季高夏季低, 差异主要由于冬季低潮带桡足类丰度的大幅增加。由分析结果可见, 季节对高潮带和中潮带丰度的影响不显著, 对低潮带季节的影响十分显著。BIOENV分析结果显示, pH、盐度、叶绿素最能解释低潮带小型底栖生物季节间群落差异。叶绿素含量可代表沉积物中微藻的丰度, 微藻可为小型底栖生物提供丰富的食物来源[28], 冬季叶绿素含量略高于夏季, 表明12月该潮间带具有较高的初级生产力, 而6月初级生产力较低。间隙水的盐度, 冬季略高于夏季, 可能是由于海南岛的降水主要集中在5~10月份[29]。夏季温度的升高, 微生物分解过程旺盛, 耗氧增多, 使得冬季低潮带的溶氧量高于夏季。Murrell 等人的研究表明桡足类对溶解氧含量十分敏感[30-33]。此外, 冬季低潮带沉积物粒径大于中高潮带, 从而更适宜于间隙生活的桡足类的生存[34]。因此, 食物丰度、溶氧量升高与沉积物粒径的变化可能是冬季低潮带桡足类丰度的大幅增加的主要原因。
低潮带与高潮带的生物量在季节间有显著差异(<0.05)。高潮带夏季生物量与丰度规律不一致, 是因为高潮带在夏季多毛类, 介形类的数量比例上升, 而且其个体干重为桡足7.5倍和14倍, 导致了生物量的差异。
3.2 丰度的潮间带变化及其与环境因子的关系
以往的研究表明[35-36], 由于中潮带有机质输入、间隙水溶解氧含量以及水分保持达到最佳的平衡点, 所以中潮带是小型底栖动物的最佳生存环境, 与高、低潮带相比, 往往支持着相对较高的底栖生物丰度和生物多样性。高潮带由于生存环境较为极端, 所以丰度较低[37]。潮间带小型底栖生物的丰度分布同样受人为扰动的影响, 扰动通常在高中潮带。Gheskiere等人的研究表明[11], 砂滩的机械清理在清除人为垃圾和海藻的同时, 也清除了中潮带的有机碎屑, 人为干扰会导致小型底栖丰度的降低。
本研究冬季潮带丰度分布趋势为, 低潮带>中潮带>高潮带, 低潮带与中高潮带有显著差异, 差异主要来源为桡足类。BIOENV分析显示溶氧量和分选系数的组合最能解释冬季潮间带小型底栖生物群落结构, 其相关系数为0.426。以往的研究结果表明, 桡足类丰度受到溶氧量[30-33], 沉积物粒径[34], 盐度[39-40]的影响。冬季大东海潮带间盐度的变化为, 高潮带>低潮带>中潮带, 除降雨外, 间隙水盐度可能主要受到地下水与海水交换的影响[18, 41-42]。冬季潮间带溶氧量表现为高潮带>低潮带>中潮带, 可能是由于高潮带沉积物被淹没时间短。因此该砂滩理化性质无法完全解释冬季潮间带的小型底栖生物丰度分布。冬季是三亚大东海砂滩的旅游旺季, 冬季人为干扰加剧可能是该砂滩丰度差异的另一原因。低潮带暴露时间较短, 受人类扰动影响较小, 人为干扰主要发生在高中潮带, 如游人在砂滩上散步、嬉戏, 加上中高潮带砂滩的清理作用, 导致中高潮带生物的丰度较低[38]。
夏季潮带间小型底栖丰度分布趋势为中高潮带大于低潮带, 特别是桡足类丰度中高潮带明显高于低潮带。与通常情况[37]有所不同。本研究结果表明, 叶绿素含量和有机质含量的组合能最好地解释夏季潮间带小型底栖生物群落的时空分布。有机质含量呈现低潮带<中高潮带, 叶绿素含量呈现低潮带<中<高潮带, 中高潮带食物更丰富。同时, 溶氧量与盐度呈现高潮带>中>低潮带, 适宜桡足类生活。由于高潮带有机质含量、间隙水和溶解氧含量达到最佳的平衡点, 形成小型底栖动物的最佳生存环境。
4 结论
本研究对三亚大东海砂质潮间带小型底栖生物及其所处的沉积环境进行了采样调查, 主要结果如下:
(1) 三亚大东海砂质潮间带共鉴定出11个小型底栖生物类群, 优势类群为海洋线虫类与桡足类;
(2) 小型底栖生物的丰度和生物量都存在显著的潮区差异和季节波动。冬季小型底栖生物的丰度和生物量都高于夏季; 冬季低潮带小型底栖生物的丰度和生物量都显著高于其他两个潮带, 而夏季则相反, 显著低于其他两个潮带。pH、偏态系数与小型底栖生物的丰度呈极显著的负相关, 分选系数、中值粒径与之呈极显著的正相关。
(3) 大东海小型底栖生物群落在季节之间差异显著。冬季潮带间差异显著, 夏季潮带间差异不显著。其中桡足类和线虫类两个类群贡献了冬季低潮带与中、高潮带群落50%以上的异质性。溶氧量和分选系数的组合最能解释冬季潮间带小型底栖生物群落的时空分布, 叶绿素含量和有机质含量的组合能最好地解释夏季潮间带小型底栖生物群落的时空分布。
[1] McLachlan A, Turner I. The interstitial environment of sandy beaches[J]. Marine Ecology, 1994, 15(3-4): 177- 212.
[2] McLachlan A, Dorvlo A. Global patterns in sandy beach macrobenthic communities[J]. Journal of Coastal Research, 2005, 21(4): 674-687.
[3] Defeo O, Mclachlan A, Schoeman D S, et al. Threats to sandy beach ecosystems: A review[J]. Estuarine Coastal and Shelf Science, 2009, 81(1): 1-12.
[4] 赵美霞, 余克服, 张乔民, 等. 应用海洋生态足迹评价三亚珊瑚礁资源的可持续利用[J]. 热带海洋学报, 2011, 30(2): 74-80. Zhao Meixia, Yu Kefu, Zhang Qiaomin, et al. Evaluating the sustainability of coral reefs in Sanya of Hainan Island using ecological footprint[J]. Journal of Tropical Oceanography, 2011, 30(2): 74-80.
[5] Giere O. Meiobenthology[M]. Berlin/Heidelberg: Springer-Verlag, 1993.
[6] Higgins R P, Thiel H. Introduction to the Study of Meiofauna[M]. Washington D C: Smithsonian institution Press, 1988.
[7] 国家海洋局. 海洋调查规范[M]. 2007. State Oceanic Administration. Specifications for Oceanographic Survey[M]. 2007.
[8] 王荣. 荧光法测定浮游植物色素计算公式的修正[J]. 海洋科学, 1986, 10(3): 1-5. Wang Rong. Correction of the formula to convert fluorescence measurement to phaeophorbide a concentration in acidification method[J]. Marine Sciences, 1986, 10(3): 1-5.
[9] 张志南, 周红, 于子山, 等. 胶州湾小型底栖生物的丰度和生物量[J]. 海洋与湖沼, 2001, 32(3): 139-147. Zhang Zhinan, Zhou Hong, Yu Zishan, et al. Abundance and biomasss of the benthic meiofauna in the northern soft-bottom of the Jiaozhou Bay[J]. Oceanologia et Limnologia Sinica, 2001, 32(2): 139-147.
[10] Widbom B. Determination of average individual dry weights and ash-free dry weights in different sieve fractions of marine meiofauna[J]. Marine Biology, 1984, 84(1): 101-108.
[11] Ganson B O. Quantitive and experimental studies of the intertidal fauna in four Swedish sandy beaches[J]. Ophelia, 1968, 5: 1-72.
[12] Raffaelli D G, Mason C F. Pollution monitoring with meiofauna: using the ratio of nematodes to copepods[J]. Marine Pollution Bulletin, 1981, 12(5): 158-163.
[13] Heip C, Vincx M, Vranken G. The ecology of marine nematodes[J]. Oceanography and Marine Biology. 1985, 23: 399-489.
[14] Clarke K, Warwick R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation[M]. Plymouth: Plymouth Marine Laboratory, 2001.
[15] Wieser W. Benthic studies in Buzzards Bay. II. The meiofauna.[J]. Limnology & Oceanography, 1960, 5(2): 121-137.
[16] Coull B C. Ecology of the Marine Meiofauna. In: Higgins R P & Thiel H. eds. Introduction to the Study of Meiofauna[M]. Washington: Smithsomian Institution Press, 1988.
[17] Somerfield P J, Rees H L, Warwick R M. Interrelationships in community structure between shallow-water marine meiofauna and macrofauna in relation to dredgings disposal[J]. Marine Ecology Progress, 1995, 127(1-3): 103-112.
[18] Hua E, Zhang Z, Zhou H, et al. Meiofauna distribution in intertidal sandy beaches along China shoreline (18°– 40°N)[J]. Journal of Ocean University of China, 2016, 15(1): 19-27.
[19] 付姗姗, 慕芳红, 杨世超, 等. 青岛沧口潮间带小型底栖生物的时空分布研究[J]. 中国海洋大学学报(自然科学版), 2012, 42(S1): 124-130. Fu Shanshan, Mu Fanghong, Yang Shichao, et al. Study on the spatial-temporal distribution of meiofauna in the intertidal zone of Cangkou, Qingdao[J]. Journal of Ocean University of China, 2012, 42(S1): 124-130.
[20] 许书会. 人为干扰对砂质潮间带小型底栖生物的影响[D]. 中国海洋大学, 2013. Xu Shuhui. Effect of human-disturbance to marine meiofauna in the sandy intertidal shores of Qingdao[D]. Ocean University of China, 2013.
[21] 李佳, 华尔, 张志南. 青岛砂质潮间带小型底栖动物分布及季节动态[J]. 应用生态学报, 2012, 23(12): 3458-3466. Li Jia, Hua Er, Zhang Zhinan. Distribution and seasonal dynamics of meiofauna in intertidal zone of Qingdao sandy beaches, Shandong Province of East China[J]. Chinese Journal of Applied Ecology, 2012, 23(12): 3458-3466.
[22] 丛冰清. 舟山砂质潮间带小型底栖生物空间分布及季节动态[D]. 青岛: 中国海洋大学, 2011. Cong Bingqing. Spatial distribution and seasonal dynamics of meiofauna in a sandy beach of Zhoushan[D]. Qingdao: Ocean University of China, 2011.
[23] 张婷. 厦门典型沙滩小型底栖动物生态学的研究[D]. 青岛: 中国海洋大学, 2011. Zhang Ting. Ecological study of meiofauna in the typical sandy beaches of Xiamen[D]. Qingdao: Ocean University of China, 2011.
[24] 韦晓慧, 慕芳红, 杨世超, 等. 青岛薛家岛砂质潮间带小型底栖生物丰度和生物量[J]. 中国海洋大学学报(自然科学版), 2013, 43(10): 60-66. Wei Xiaohui, Mu Fanghong, Yang Shichao, et al. Study on the abundance and biomass of meiofauna in the intertidal sands of Xuejiadao in Qingdao[J]. Journal of Ocean University of China, 2013, 43(10): 60-66.
[25] 林岿璇, 张志南, 韩洁. 南麂列岛海洋自然保护区潮间带小型生物初步研究[J]. 中国海洋大学学报(自然科学版), 2003, 33(2): 219-225. Lin Kuixuan, Zhang Zhinan, Han Jie. A preliminary study on the meiofauna in the intertidal zone in Nanji Islands marine reserve[J]. Journal of Ocean University of Qingdao, 2003, 33(2): 219-225.
[26] 张志南. 秦皇岛砂滩海洋线虫的数量研究[J]. 青岛海洋大学学报(自然科学版), 1991, 21(1): 63-75. Zhang Zhinan. Marine Nematodes on four intertidal sandy beaches at the Qin Huang Dao Bay, North China[J]. Journal of Ocean University of Qingdao, 1991, 21(1): 63-75.
[27] 曹英昆, 李垚垚, 霸婉玉, 等. 秦皇岛海水浴场海滩小型底栖动物丰度的研究[J]. 北京联合大学学报, 2018, 32(2): 45-50. Cao Yingkun, Li Yaoyao, Ba Wanyu, et al. Study on abundance of meiofauna on the bathing beach of Qinhuangdao[J]. Journal of Beijing Union University, 2018, 32(2): 45-50.
[28] Armonies W, Reise K. Faunal diversity across a sandy shore[J]. Marine Ecology Progress Series, 2000, 196: 49-57.
[29] 王胜, 吴坤悌, 陈明. 1961年以来海南岛降水变化趋势分析[J]. 气象研究与应用, 2006, 27(1): 24-27. Wang Sheng, Wu Kunti, Chen Ming. Analysis for precipitation change trend of Hainan Island since 1961[J]. Journal of Meteorological Research and Application, 2006, 27(1): 24-27.
[30] Murrell M C, Fleeger J W. Meiofauna abundance on the Gulf of Mexico continental shelf affected by hypoxia[J]. Continental Shelf Research, 1989, 9(12): 1049-1062.
[31] Levin L A, Huggett C L, Wishner K F. Control of deep- sea benthic community structure by oxygen and organic-matter gradients in the eastern Pacific Ocean[J]. Journal of Marine Research, 1991, 49(4): 763-800.
[32] Neira C, Sellanes J, Levin L A, et al. Meiofaunal distributions on the Peru margin: relationship to oxygen and organic matter availability[J]. Deep Sea Research Part I Oceanographic Research Papers, 2001, 48(11): 2453-2472.
[33] Hua E, Zhang Z, Zhang Y. Meiofauna distributions at the oxygen minimum zone in Changjiang (Yangtze) River Estuary waters[J]. Acta Oceanologica Sinica, 2006, 25(5): 120-134.
[34] McLachlan A, Bate G C. Sandy Beach Ecology[M]. Springer Netherlands, 1983.
[35] Gheskiere T, Hoste E, Vanaverbeke J, et al. Horizontal zonation patterns and feeding structure of marine nematode assemblages on a macrotidal, ultra-dissipative sandy beach De Panne, Belgium[J]. Journal of Sea Research, 2004, 52: 211-226.
[36] McLachlan A, Brown, A C. The Ecology of Sandy Shores[M]. Academic Press, 2006: 372.
[37] Knox G A. The Ecology of Seashores[M]. CRC Press, 2001: 557.
[38] Gheskiere T, Vincx M, Weslawski J M, et al. Meiofauna as descriptor of tourism-induced changes at sandy beaches[J]. Marine Environmental Research, 2005, 60(2): 245-265.
[39] Ingole B S, Parulekar A H, Parulekar A H. Role of Salinity in structuring the intertidal meiofauna of a tropical estuarine beach: Field evidence[J]. Indian Journal of Marine sciences, 1998, 27: 356-361.
[40] Priyalakshmi C, Menon N R. Ecology of interstitial faunal assemblage from the beaches along the coast of Kerala, India[J/OL]. International Journal of Oceanography, 2014, (2014): Article ID 284979, 1-9. https: //doi. org/10.1155/2014/284979.
[41] Priyalakshmi G, Menon N R. Ecology of interstitial faunal assemblage from the beaches along the coast of Kerala, India[J]. International Journal of Oceanography, 2014: 1-9.
[42] Cai L Z, Fu S J, Yang J, et al. Distribution of meiofaunal abundance in relation to environmental factors in Beibu Gulf, Southe China Sea[J]. Acta Oceanologica Sinica, 2012, 31(6): 92-103.
The spatio–temporal distribution of meiofauna in Dadonghai beach (Sanya, China) and its influcing factors
LUO Tian, LIANG Chen-zi, SUN Yan, YANG Li-li, LI Ya-nan, ZANG Yu, MU Fang-hong
(College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China)
A seasonal survey of intertidal meiofauna and its sedimentary environment in the sandy beach of Dadonghai, Sanya was conducted in 2015 and 2016. The spatio–temporal distribution of meiofaunal abundance, biomass and the community structure were investigated, and the influencing environmental factors were analyzed. Total 11 meiofauna groups were identified. The most dominant groups were nematodes and copepods, accounting for 83.13% of the total abundance. The average meiofaunal abundance was (595.77 ± 442.72) ind·10 cm−2. The average meiofaunal abundance in December 2015 was (768.70 ± 533.22) ind 10 cm−2, and that in June 2016 was (422.85 ± 236.42) ind 10 cm−2. The meiofaunal abundance varied spatially and temporally. pH and skew coefficient (SKΦ) are negatively correlated with meiofaunal abundance, whereas sorting coefficient (QDΦ) and median grain size (MDΦ) are positively correlated with meiofaunal abundance. The meiofaunal abundance tends to be greater in winter than in summer. Food, dissolved oxygen, and sediment particle size were the main reasons for the increase of copepods in winter. According to two-way ANOVA and ANOSIM, the abundance and community structure of meiofauna reflected a significant difference among different tides in winter. According to SIMPER analysis, this difference can be mostly attributed to nematodes and copepods. According to BIOENV analysis, the combination of dissolved oxygen and QDΦcould best explain the situation, which could result in human interference and mechanical cleaning that occurs in high and middle tides. In summer, no significant difference among different tides can be observed. The combination of pH, Chland organic matter content could best explain the meiofauna distribution in summer.The results can provide basic data for further research on interstitial meiofauna, together with scientific basis for the protection, exploitation, and utilization of sandy intertidal zones under the background of global climate change.
meiofauna; spatio–temporal distribution; abundance; community structure; Dadonghai Beach; Sanya
Jan. 19, 2019
[National Natural Science Foundation of China, No. 41576153, No. 41976100]
Q178.531
A
1000-3096(2020)02-0036-09
10.11759/hykx20190119001
2019-01-19;
2019-03-21
国家自然科学基金项目(41576153; 41976100)
骆添(1993-), 女, 江西九江人, 硕士研究生, 电话: 17863927463, E-mail: 1748157991@ qq.com; 慕芳红(1972-),通信作者, 副教授, 博士, 研究方向: 底栖生态学, E-mail: fanghongmu@ouc. edu.cn
(本文编辑: 杨 悦 李晓燕)
猜你喜欢
铁道建筑技术(2022年6期)2022-07-13
河北地质(2021年1期)2021-07-21
湘潮(上半月)(2021年4期)2021-07-20
中国水产(2021年1期)2021-03-01
黑龙江水利科技(2020年8期)2021-01-21
海洋信息技术与应用(2020年4期)2021-01-18
矿产勘查(2020年5期)2020-12-25
海洋通报(2020年3期)2020-10-20
安徽农业科学(2018年21期)2018-05-14
体育世界·扣篮(2007年12期)2007-11-30
