
不同品种烤烟衰老期碳氮代谢及相关基因表达的差异分析
2020-03-03 03:08张小全蔡文龙阚洪赢薛冰洁贾振宇王帅武云杰
中国烟草学报 2020年6期
张小全,蔡文龙,阚洪赢,薛冰洁,贾振宇,王帅,武云杰
河南农业大学烟草学院,郑州市农业路63号 450002
烟草以收获叶片为目的,烟叶成熟的过程就是逐渐衰老的过程,也是烟叶品质形成最关键的时期[1-2]。烟叶的碳氮代谢是烟叶最基础的物质代谢,与施氮量、品种等因素密切相关[3-4]。碳代谢包括无机碳的光合固定代谢、单糖的转化以及淀粉的积累等过程[5]。颗粒结合型淀粉合成酶(GBSS)参与直链淀粉的合成[6],淀粉分支酶(SBE)是合成支链淀粉的关键酶[7],两个关键酶的活性,是反应碳代谢强度的重要指标。氮代谢包括氮素的吸收、同化、转运、利用、调节等过程[5]。硝酸还原酶(NR)和谷氨酸脱氢酶(GDH)分别在氮素的吸收和转运方面具有重要作用[8-9],谷氨酰胺合成酶(GS)是控制铵离子转移和再同化的关键酶[10-11]。大量研究发现,烟叶的成熟衰老过程和碳氮代谢的变化紧密相关。在烟叶生长和成熟过程中,以碳水化合物为主的碳积累代谢逐渐增强,而碳的固定和转化代谢逐渐减弱[5],随着碳代谢达到一定程度,淀粉的降解大于积累,烟叶成熟度增加,烟叶品质提升[2]。随着烟叶生长和衰老程度加深,总氮和蛋白质等氮素营养物质的合成速度下降,降解速度增加[12]。在烟叶衰老期,蛋白水解酶被逐渐释放到细胞质,促进蛋白质的降解[13],蛋白质的降解使色素蛋白复合体分解,光合碳同化能力下降,并进一步促进质体色素的分解[14],因而,叶绿素和类胡萝卜素对衰老十分敏感,其含量下降是烟叶衰老的标志[15]。铵离子是烟叶衰老后期氮素转移和再利用的氮代谢中间体[16-17],胞质谷氨酰胺合成酶(GS1)定位于胞液中,其在叶片衰老过程中调控氮源的转移及再利用,而叶绿体谷氨酰胺合成酶(GS2)定位于叶绿体中,主要作用是氨同化[18]。周健飞研究发现,烤烟叶片叶肉中GS2 蛋白亚基含量随着叶龄的增长逐渐下降,GS2 活性与质外体铵浓度和氨气挥发量呈负相关[19]。而对不同烤烟品种衰老特性与碳氮代谢特征间的关系还缺乏系统的研究。
本研究选取4 个田间成熟落黄表现不同的烤烟品种,对不同叶龄中部叶中的质体色素、碳氮代谢指标及相关基因表达量的变化差异进行了研究,以期从品种角度解读烤烟成熟衰老特性差异与碳氮代谢特性的关系,为调控烟叶衰老进程、选育适宜衰老特性的品种提供理论依据。
1 材料与方法
1.1 材料
以田间成熟落黄表现不同的烤烟品种豫烟10号、云烟87、K326 和NC89 为试验材料。4 个品种在相同的栽培管理条件下,中部叶的发育进程基本一致,生理成熟期都是在叶龄70 d 左右。种子由河南农业大学烟草育种实验室提供。
1.2 试验设计
试验于2018 年在河南农业大学许昌校区进行。供试土壤为肥力一致的轻壤土,基本理化性状为有机质10.77 g/kg、全氮0.96 g/kg、碱解氮48.7 mg/kg,速效磷51.8 mg/kg,速效钾103.7 mg/kg,pH 7.56。试验采用完全随机区组设计,3 次重复,行距120 cm,株距50 cm。5 月5 日进行移栽,使用烟草专用复合肥(10-12-18)、重过磷酸钙和硫酸钾,纯氮用量45 kg/hm2,N : P2O5 : K2O=1 : 2 : 3,基肥与追肥比例为7 : 3,追肥于6 月5 日施入,田间管理参照优质烤烟生产技术进行,打顶后单株留叶20 片。参照文献[20]的方法进行取样和相关指标的测定。每个品种分别选取整齐一致的40 至50 株烟,在叶长1.5 cm时标记为叶龄1 d,在叶龄30~70 d,每隔10 d,每个品种每次从3~5 株第10~12 片叶位(自下向上数)上取适龄叶片,同一样品取叶片中部后充分剪片混匀分3 份保存,分别用于质体色素含量测定,相关基因表达量测定和杀青后相关化学成分测定。
1.3 测定项目及方法
采用分光光度法测定叶绿素、类胡萝卜素含量[21]。总氮、总碳用全自动CN分析仪(Vario MAX CN,德国)测定;还原糖、总糖、淀粉含量采用AA3 型连续流动化学分析仪(德国 BRAN+LUEBBE 公司生产)测定[22]。
参照文献[20]的方法计算目的基因的相对表达量,并绘制相关图形。以烟草L25 基因为内参基因,引物序列见表1。
1.4 数据处理与分析
采用Excel 2016 和SPSS21.0 软件处理和分析试验数据及绘制图形,用Duncan 法进行显著性检验。
2 结果与分析
2.1 不同品种叶片衰老过程中质体色素含量变化
4 个烤烟品种中部叶,叶龄30 d 后,叶绿素含量和类胡萝卜素含量均随着叶龄的增大逐渐降低(表2)。不同品种间降低的幅度有较大的差异。豫烟10 号叶绿素含量在叶龄70 d 时显著低于其他品种(P<0.05),NC89 从叶龄40 d 开始,叶绿素含量均高于其他品种。NC89 的类胡萝卜素含量在各时期均较高。从叶龄30~70 d,豫烟10 号、云烟87、K326 和NC89 的叶绿素含量降幅分别为53.52%、46.43%、50.19%、43.44%,类胡萝卜素含量的降幅分别为48.83%、39.13%、44.40%、40.00%。质体色素含量的变化和4 个品种中部叶的田间落黄表现相一致。
2.2 不同品种叶片衰老过程中类胡萝卜素降解基因表达量变化
不同品种叶片成熟衰老过程中,类胡萝卜素裂解双加氧酶基因(CCD)表达量逐渐升高(图1),品种间有较大的差异。豫烟10 号除叶龄60 d 外其他时期均极显著高于其他品种。叶龄40 d 时云烟87 和K326 较上一时期有较大增幅,NC89 的表达量在叶龄40 d、50 d 和70 d 均低于其他3 个品种,成熟后期(叶龄70 d)极显著地低于其他品种(P<0.01)。云烟87 和K326 在叶龄70 d 的表达量居中。不同品种的CCD 表达量与其叶片类胡萝卜素含量高低趋势相反,CCD 表达量低,类胡萝卜素含量相对较高。
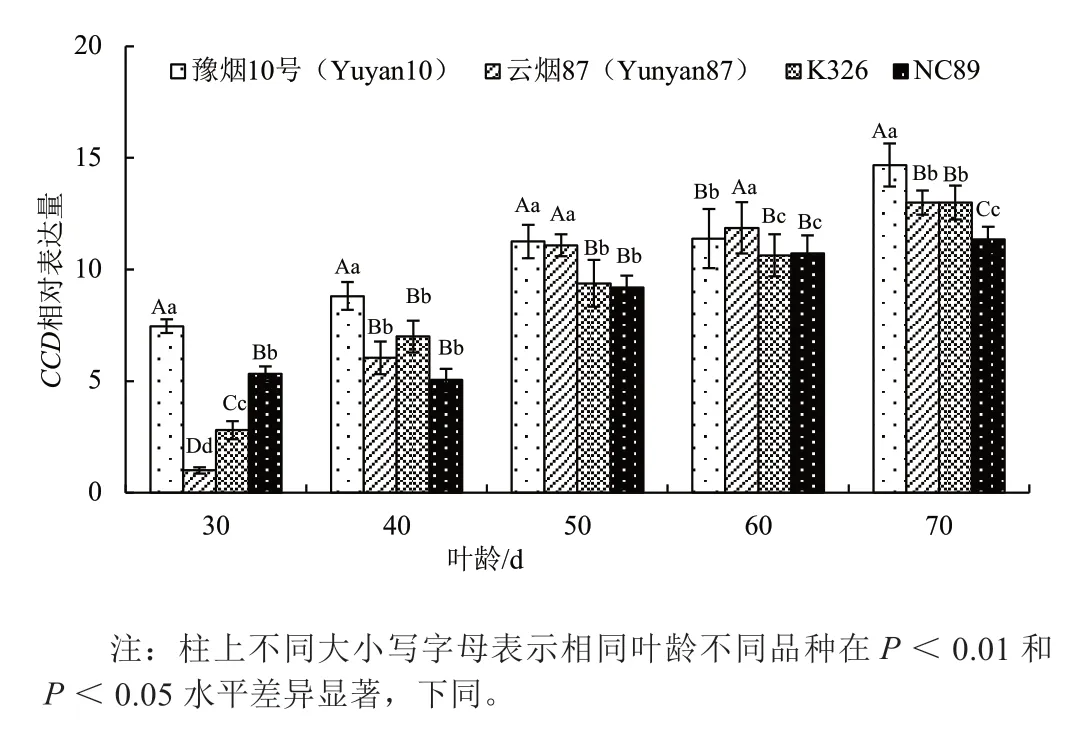
图1 不同品种叶片衰老过程中CCD 基因相对表达量的变化Fig.1 The dynamic changes of CCD gene relative expression in leaves of different flue-tobacco varieties during senescence phase
2.3 不同品种叶片衰老过程中碳代谢相关指标变化
2.3.1 叶片中淀粉、还原糖、总糖和总碳含量变化
中部叶片衰老过程中,除NC89 外,其他3 个烤烟品种的淀粉含量在40 d 时达到峰值(图2A),在叶龄40~50 d 间,四个品种中豫烟10 号的淀粉含量降低量最大,NC89 的淀粉含量变化较小。从叶龄40~70 d,豫烟10 号的淀粉含量降幅最大(38.65%),NC89 最小(27.94%),云烟87 和K326 的淀粉含量变化趋势一致。还原糖和总糖含量在衰老过程中表现出先升高后下降的趋势(图2B、2C),豫烟10 号的还原糖含量在叶龄50 d 时达到峰值,另外3个品种均在叶龄60 d 时达到峰值。叶龄60~70 d,豫烟10 号、K326、云烟87 和NC89 还原糖含量分别下降16.14%、14.80%、12.52%和10.00%。总糖的含量均在叶龄50 d 时达到峰值,叶龄50 d 至70 d,豫烟10 号、K326、云烟87 和NC89 的总糖含量分别下降22.49%、17.72%、20.00%和11.50%。叶片总碳含量在烟叶在衰老过程中维持在46%~50%之间(图2D),整体表现出先升高后降低的规律,在叶龄40 d 或50 d 时达到峰值后又缓慢下降;叶龄50~60 d 期间,豫烟10 号总碳均极显著低于其他品种(P <0.01),云烟87 和K326 显著低于NC89(P <0.05);NC89 在整个生长期间均保持较高的总碳含量,叶龄70 d 时,4 个品种间存在显著的差异(P <0.05),表现为NC89>云烟87>豫烟10 号>K326。

图2 不同品种叶片衰老过程中碳代谢相关物质含量的变化Fig. 2 The dynamic changes of carbon metabolizes nutrients in leaves of different flue-tobacco varieties during leaf senescence phase
2.3.2 叶片中碳代谢相关基因表达量变化
对叶片中碳代谢相关基因颗粒结合型淀粉合成酶基因(GBSS1)(图3A)和淀粉分支酶基因(SBE)(图3B)的表达量变化分析发现,两个淀粉合成关键基因在4 个品种中的表达量均表现出先升高后下降的趋势,且品种间有较大的差异,与淀粉含量在衰老进程中的表现相一致。豫烟10 号两个基因的表达量在叶龄40 d 前显著地高于NC89(P<0.05),NC89两个基因的表达量在叶龄50d 后极显著地高于其他3个品种(P<0.01)。在叶龄 60 d 和 70 d 时 NC89 的GBSS1 表达量分别是豫烟10 号的2.40 倍和2.05 倍。云烟87 和K326 两个基因的表达量在叶龄50 d 前差异较大,叶龄50 d 后表达量变化趋势基本一致。
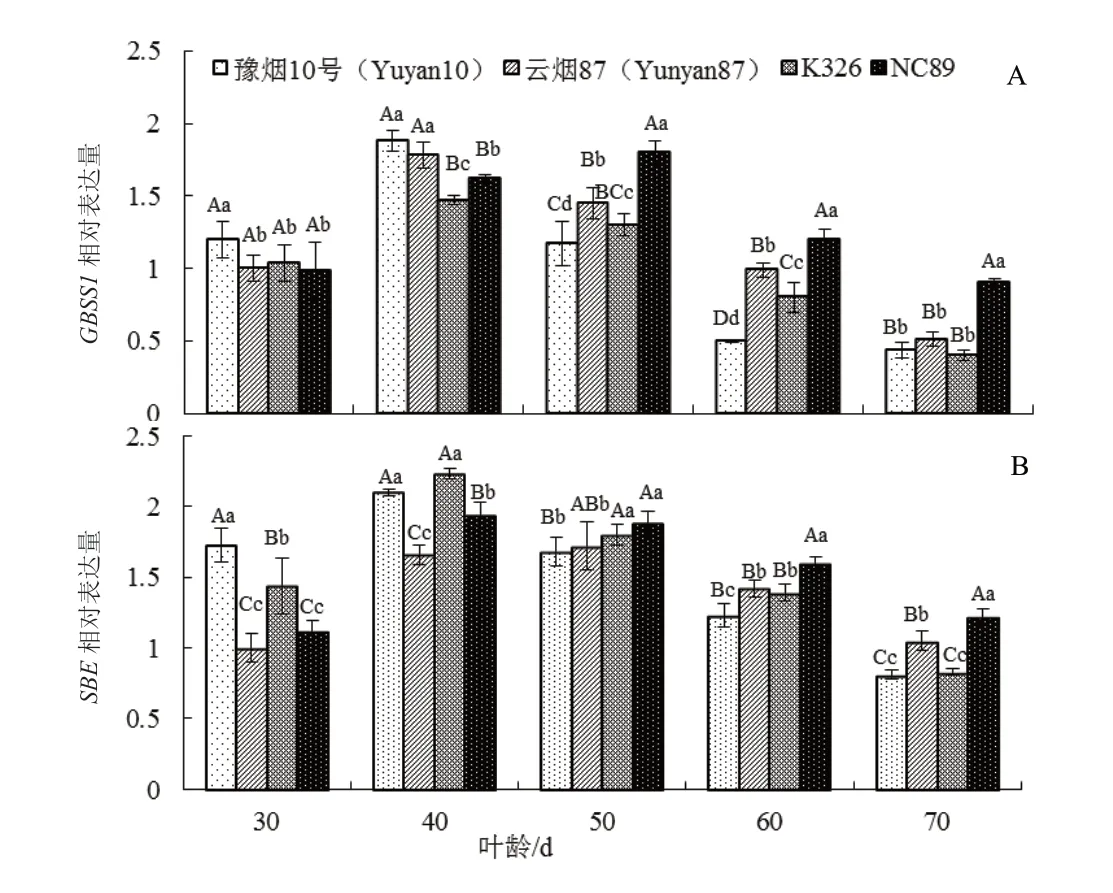
图3 叶片衰老过程中碳代谢相关基因相对表达量的变化Fig. 3 Changes of relative expression of carbon metabolism related genes during leaf senescence phase
2.4 不同品种叶片衰老过程中氮代谢指标变化
2.4.1 叶片中烟碱和总氮含量变化
在相同的栽培管理条件下,叶片成熟衰老过程中,不同品种烟碱和总氮含量的变化具有较大差异(图4)。NC89 叶片的烟碱含量在成熟衰老过程中均极显著地高于其他品种(P<0.01),在叶龄60 d和70 d 分别是豫烟10 号含量的1.96 倍和2.10 倍(图4 A)。叶龄30~50 d,豫烟10 号、云烟87 和K326的烟碱含量差异较小,叶龄60~70 d,豫烟10 号的烟碱含量显著低于云烟87 和K326。叶片成熟衰老过程中,总氮含量除NC89 从叶龄30~40 d 有小幅上升外,其余品种均表现为下降趋势(图4B);叶龄40~50 d 时豫烟10 号降幅最大,并在叶龄60 d 时显著低于其他品种,从叶龄30~60 d 豫烟10 号的总氮含量下降27.95%,NC89 仅8.91%。叶片成熟后期(叶龄60~70 d 时),NC89 的总氮含量下降较快,并在叶龄70 d 时显著低于其他3 个品种。
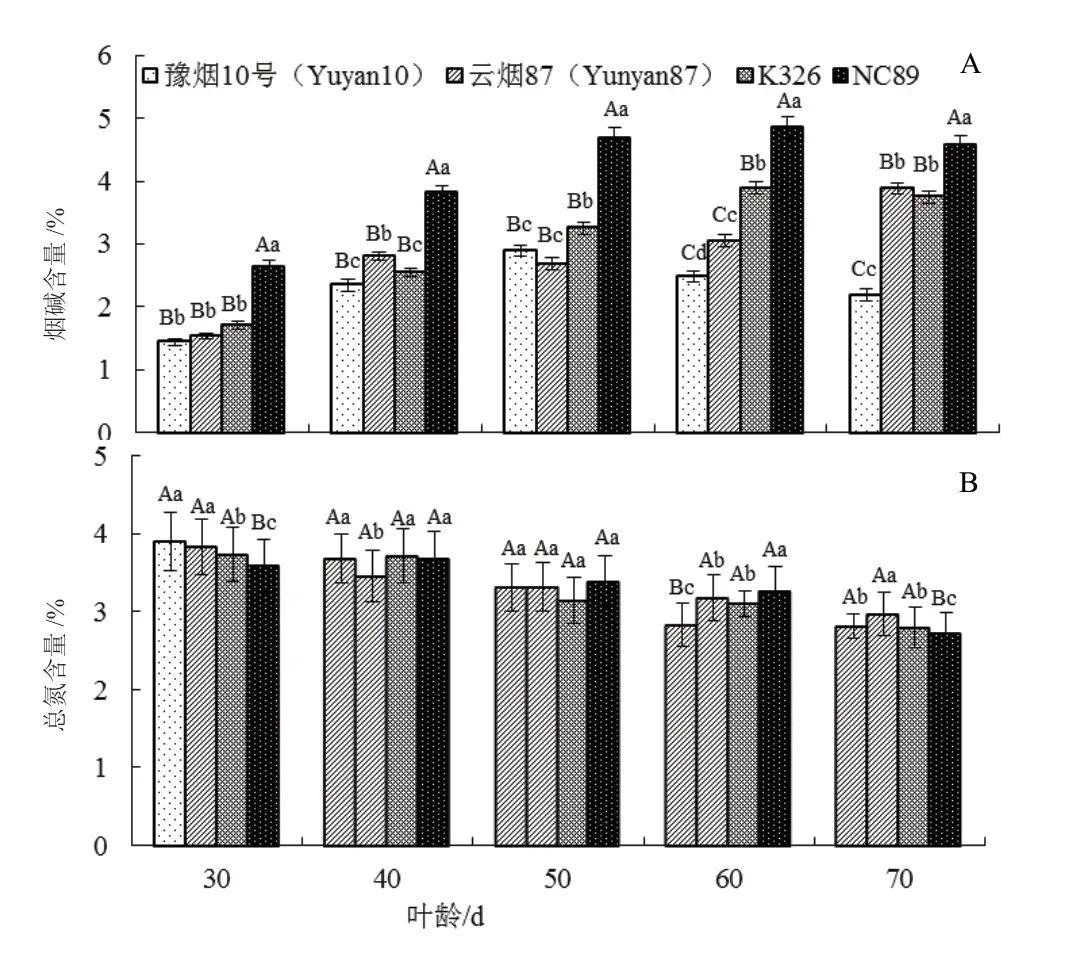
图4 不同品种叶片衰老过程中烟碱和总氮含量的变化Fig. 4 The dynamic changes of nicotine and total nitrogen contents in leaves of different flue-tobacco varieties during leaf senescence phase
2.4.2 叶片中氮代谢相关基因表达量变化
在烟草叶片成熟衰老过程中,调控氮素转移和再利用的GS1-3 基因相对表达量呈上升趋势(图5A),调控氨同化的GS2 基因相对表达量呈下降趋势(图5B),不同品种变化规律基本一致,但品种间存在较大的差异。豫烟10 号的GS1-3 基因表达量在整个生长衰老期显著高于其他品种(P<0.05)。叶龄50 d 之后,NC89 品种GS1-3 的表达量均显著低于其他品种(P<0.05),云烟87 和K326 间的表达量差异较小。NC89 品种GS2 的基因表达量在整个生长衰老期均极显著地高于豫烟10 号(P<0.01),云烟87 和K326 的GS2 基因表达量的变化幅度居中。说明4 个品种同化利用细胞中各种氮源的方式和能力具有较大差异。
在烟叶成熟衰老过程中,叶片中控制氮素吸收的硝酸还原酶(NR)基因相对表达量逐渐降低(图5C),豫烟10 号NR 基因的表达量叶龄30 d 时极显著的高于其他品种(P<0.01),在叶龄50 d 之后均显著低于其他品种(P<0.05),说明在叶片成熟过程中其氮素同化能力下降较大;NC89 在叶龄60 d 后,NR 基因表达量极显著的高于其他品种(P<0.01),说明NC89 成熟后期仍保持较高的氮素同化能力。除NC89 外,其他品种调控氮素转运的谷氨酸脱氢酶基因(GDH1)相对表达量均在叶龄50 d 时达到峰值(图5D),NC89 在叶龄60 d 时达到峰值。豫烟10 号和K326 的GDH1 表达量在叶龄50 d 之后表达量迅速降低,NC89 和云烟87 还保持较高的表达量,说明NC89 和云烟87 叶片成熟后期仍具有较高的氮素转运能力。

图5 叶片衰老过程中氮代谢相关基因相对表达量的变化Fig. 5 Changes of relative expression of nitrogen metabolism related genes in leaves during leaf senescence
2.5 不同品种叶片衰老过程中碳氮比变化
在烟叶的衰老过程中,烟叶碳氮比(C/N)呈逐渐上升的趋势,4 个品种的碳氮比都在适宜的范围内(表3)。整体上总碳含量变化不大,C/N 的变化主要归因于总氮含量降低。叶龄50~60 d,豫烟10 号的 C/N 增幅最大,K326 最小;叶龄 60~70 d,NC89 增幅最大,豫烟10 号最小,云烟87 和K326居中。

表3 不同烤烟品种叶片衰老过程中碳氮比的变化Tab. 3 The dynamic changes of C/N ratio in leaves of different flue-cured tobacco varieties during leaf senescence phase
3 讨论
碳氮代谢是烟草生长发育基本的代谢过程[23-24],不同成熟衰老特性的烟草品种在碳氮代谢过程有较大的差异。本研究表明,成熟衰老较慢的品种NC89 的淀粉和水溶性糖含量的峰值滞后于另外3 个品种,成熟衰老较快的品种豫烟10 号叶龄50 d 后,淀粉合成基因GBSS1 表达量大幅下降,淀粉含量降低,而NC89 的淀粉合成基因GBSS1 表达量和淀粉含量的降幅较小,可能跟豫烟10 号的叶绿素和类胡萝卜素降解较快较早,影响光合作用有关。张生杰等[25]对不同品种烤烟叶片衰老和光合特性的研究证实了这一点。豫烟10 号参与氮素转移的GS1-3 基因表达量高,与氮同化相关的GS2、GDH1 和NR 基因表达量低,氮素转移量大,同化量少,含氮物质快速降解,衰老速度加快,可能是其衰老较快的原因,周健飞等对烟草成熟度的研究中也有类似的现象[19]。可见,调控烟叶衰老落黄的关键是在烟叶成熟过程中,控制氮素的同化,增加氮素转移[26]。
烟草的碳氮代谢平衡与其生长发育和烟叶品质紧密相关。烟草的碳氮代谢受品种、土壤类型、施肥量、施肥种类、水肥管理和生长环境等众多因素的影响[27-28],不同研究者从碳氮代谢相关酶,代谢关键基因的表达等方面对不同条件下烟草的碳氮代谢规律进行了研究[29-30]。本研究运用全自动CN 分析仪,对不同品种烟叶的衰老过程中碳氮比变化进行了分析,发现中部烟叶C/N 呈逐渐上升,但整体上总碳含量变化不大,C/N 变化主要归因于总氮含量变化,可能跟本研究是在相同的水肥管理条件下进行,都能正常成熟落黄,4 个品种的C/N 变化主要跟其氮代谢的差异相关。
本研究还发现,不同品种的衰老特性除了与其碳氮代谢特征相关外,还与其遗传基础紧密相关。云烟87 是由K326 和云烟2 号杂交选育的[31],豫烟10 号是由云烟87 和农大202 杂交选育的[32],云烟87 和K326 具有相似的碳氮代谢和衰老特性,豫烟10 号则与其亲本农大202 易落黄易烘烤的特性相似。NC89的碳氮代谢和衰老特性,可能与其遗传基础特殊有关。对NC89 碳氮代谢和衰老特性遗传基础的深入解读,将有助于培育适宜衰老落黄特性的优良品种。对这几个材料叶片发育过程中衰老标志基因表达变化、激素代谢的变化、不同代谢酶类分析等方面的研究也证实了类似的规律[4,12,20,25-26]。不同烟草品种碳氮代谢和衰老特性差异的原因还可以借鉴水稻等其他作物的研究成果,从根系发育特性差异[33]、营养吸收利用差异[33]、遗传基因差异[34]等方面进行深入的研究。
4 结论
烤烟品种的衰老特性差异与碳氮代谢有关。与NC89 相比,豫烟10 号中部叶衰老速度快,与其叶龄50 d 后淀粉合成基因GBSS1 表达量下降速度快,淀粉含量降低较快的碳代谢特征,以及叶片成熟衰老过程中氮素转移基因GS1-3 表达量高,氮同化相关的GS2、GDH1 和NR 基因表达量低,氮素转移量大,同化量少,含氮物质快速降解的氮代谢特征相关。
猜你喜欢
麦类作物学报(2021年12期)2022-01-08
中国土壤与肥料(2021年5期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
农业技术与装备(2020年10期)2020-11-03
云南农业(2020年5期)2020-06-12
塔里木大学学报(2019年4期)2020-01-03
种业导刊(2019年9期)2019-10-21
农民致富之友(2017年7期)2017-04-27
农民致富之友(2017年2期)2017-03-23
诗歌月刊(2014年3期)2015-04-14
