
南美白对虾病原性豚鼠气单胞菌的分离与药敏特性
2020-02-24 07:49曾宪凯曹海鹏穆春华
水产养殖 2020年1期
曾宪凯,曹海鹏,穆春华
(1.上海市青浦区练塘镇农业综合服务中心,上海鹏豪水产专业合作社,上海 201715;2.上海海洋大学农业部淡水水产种质资源重点实验室,上海市水产养殖工程技术研究中心,上海高校知识服务平台上海海洋大学水产动物遗传育种中心(ZF1206),上海 201306)
南美白对虾(Penaeus vannamei)是我国重要特种水产养殖品种,在浙江、广西、海南、福建、山东、珠三角、长三角、京津唐等地区大面积养殖[1],是我国养殖产量最高的对虾养殖品种。2017年,我国南美白对虾养殖产量高达160多万吨[2]。然而,细菌性病害对南美白对虾养殖产生了严重危害,其病原菌种类广、繁殖速度快、致病力强、发病率高,通常引起南美白对虾内脏组织出现不同程度损伤而使其大规模死亡。其中,以“附肢或游泳足变黄”为典型病症的黄腿病是南美白对虾养殖过程中的常见病之一[3-4],但从现有文献资料来看,南美白对虾黄腿病的研究基础薄弱,鲜见南美白对虾黄腿病病原菌豚鼠气单胞菌及其防控药物等方面的详细研究报道,这在一定程度上影响了我国对南美白对虾黄腿病的有效防控。因此,加强黄腿病研究对促进南美白对虾养殖业健康发展具有重要意义。2018年7月,上海市青浦区一养殖场的南美白对虾出现了黄腿病,病死率达到了60%以上,在上海南美白对虾养殖区发生实为罕见。鉴此,该试验通过对南美白对虾黄腿病病原菌的分离鉴定与药敏特性研究,以期为确定南美白对虾黄腿病的病因与药物防控提供理论资料。
1 材料与方法
1.1 材料
自然发病南美白对虾,16尾,平均体质量5.12 g,取自上海市青浦区自然发病养殖池;健康南美白对虾,60尾,大小均匀,体质健壮,平均体质量8.72 g,由浙江萧山某养殖场提供;API ID32GN细菌生化鉴定试剂条,由法国某公司生产;药敏纸片,由杭州某微生物试剂有限公司生产。
1.2 方法
1.2.1 病原菌的分离 采用75%酒精对具有典型发病症状的濒死南美白对虾进行表面消毒后,参照Cao等[4]的方法常规无菌操作取其肝胰腺组织,划线普通营养琼脂平板进行病原菌的分离,30℃恒温培养24~32 h后纯化分离菌株,并转接至普通营养琼脂斜面培养基上于30℃恒温培养32 h,4℃冰箱保存备用。
1.2.2 分离菌株的人工回归感染试验 试验包括3个试验组和1个对照组,每组各15尾健康南美白对虾。试验前用无菌生理盐水分别洗下普通营养琼脂斜面培养基上各分离菌株的菌苔制成菌浓度为1.0×106CFU/mL的菌悬液。试验组南美白对虾分别在靠近尾扇第3腹节处[5]注射浓度为1.0×106CFU/mL的分离菌株菌悬液,每尾注射0.1 mL;对照组注射相同剂量的无菌生理盐水。试验周期为6 d,试验期间水温控制在28℃,每天记录试验南美白对虾的死亡数量,观察其病症,并将死虾及时捞出进行病原菌的再分离,确定其与原病原菌株的一致性。
1.2.3 病原菌的鉴定 参照曹海鹏等[6]的方法,用API ID32GN细菌鉴定系统对病原菌株进行生理生化鉴定,并委托上海某生物科技有限公司对病原菌株进行PCR扩增与测序后,采用邻接法构建系统发育树,通过16S rRNA序列分析对病原菌株进行分子鉴定。
1.2.4 病原菌药敏特性的测定 参照纸片扩散法[7]测定病原菌株的药敏特性,并根据杭州滨河微生物试剂有限公司的《纸片法药敏试验抑菌圈直径判断标准》,判定病原菌株对抗生素的敏感性。
2 结果
2.1 病原菌的分离
从自然发病的南美白对虾的肝胰腺组织中分离到3株疑似病原菌株,暂命名为QPX1、QPX2、QPX3。人工回归感染试验结果(表1)表明,仅菌株QPX1感染南美白对虾后南美白对虾出现了死亡,试验南美白对虾的病死率达到了100%,而且试验南美白对虾的病症与自然病症基本相同,并从人工感染的病虾肝胰腺中又可再次分离到与原菌株生理生化特性一致的菌株,由此判定菌株QPX1是引起该病的病原菌。
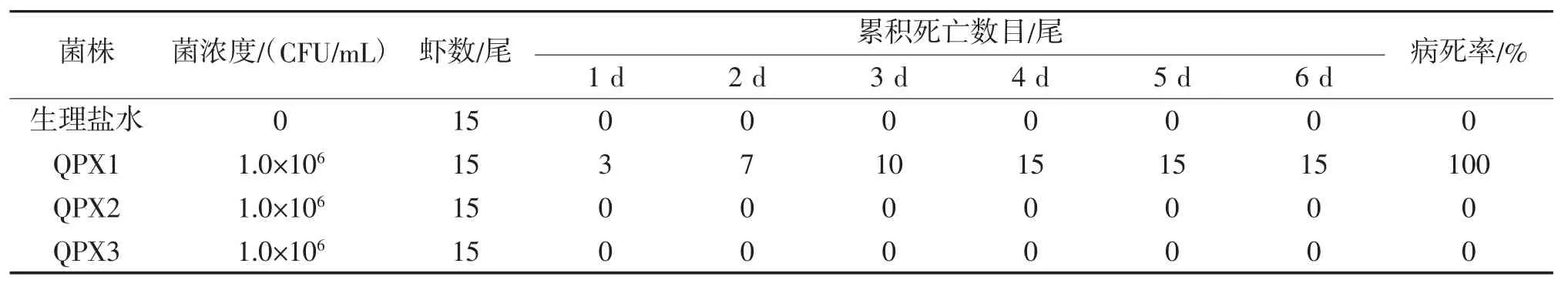
表1 分离菌株对南美白对虾人工回归感染试验结果
2.2 病原菌的鉴定
菌株QPX1的生理生化鉴定结果表明(表2),菌株QPX1为豚鼠气单胞菌(Aeromonas caviae),与豚鼠气单胞菌对照菌株[6,8]生理生化特性有96.9%的一致性。此外,通过NCBI网站中的BLASTN软件将菌株QPX1的16S rRNA基因序列(GenBank收录号:MK652096)与GenBank中已知菌株的16S rRNA基因序列进行了同源性比较(表3),发现菌株QPX1的16 S rRNA基因序列与GenBank中豚鼠气单胞菌菌株的16 S rRNA序列自然聚类,同源性达到了99%~100%。通过邻接法构建的基于菌株QPX1 16S rRNA基因序列的系统发育分析(图1)进一步表明,菌株QPX1与豚鼠气单胞菌菌株J2(GenBank收录号:KP262414)的亲缘关系最近。结合生理生化鉴定以及16S rRNA基因序列分析的结果,判定QPX1为豚鼠气单胞菌。
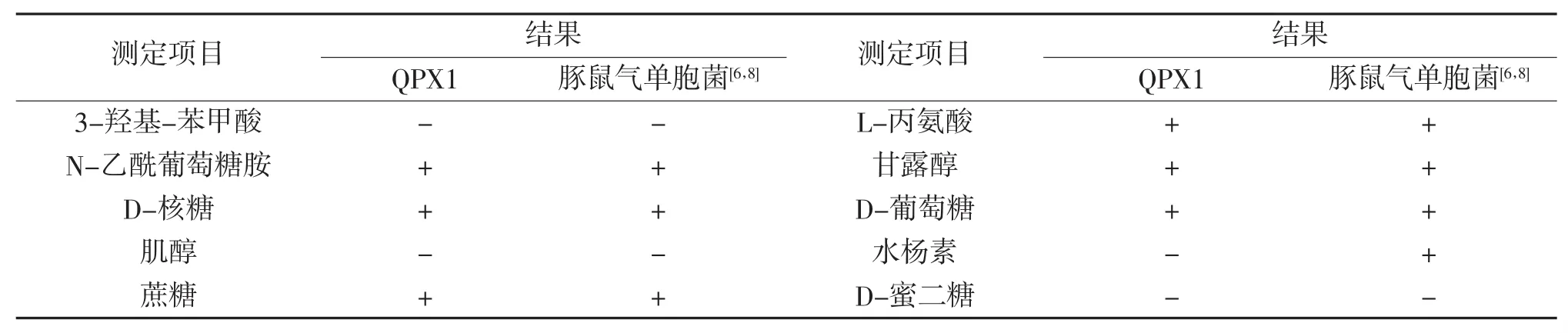
表2 菌株QPX1的生理生化特征

注:+,阳性;-,阴性;d,存在种间差异。
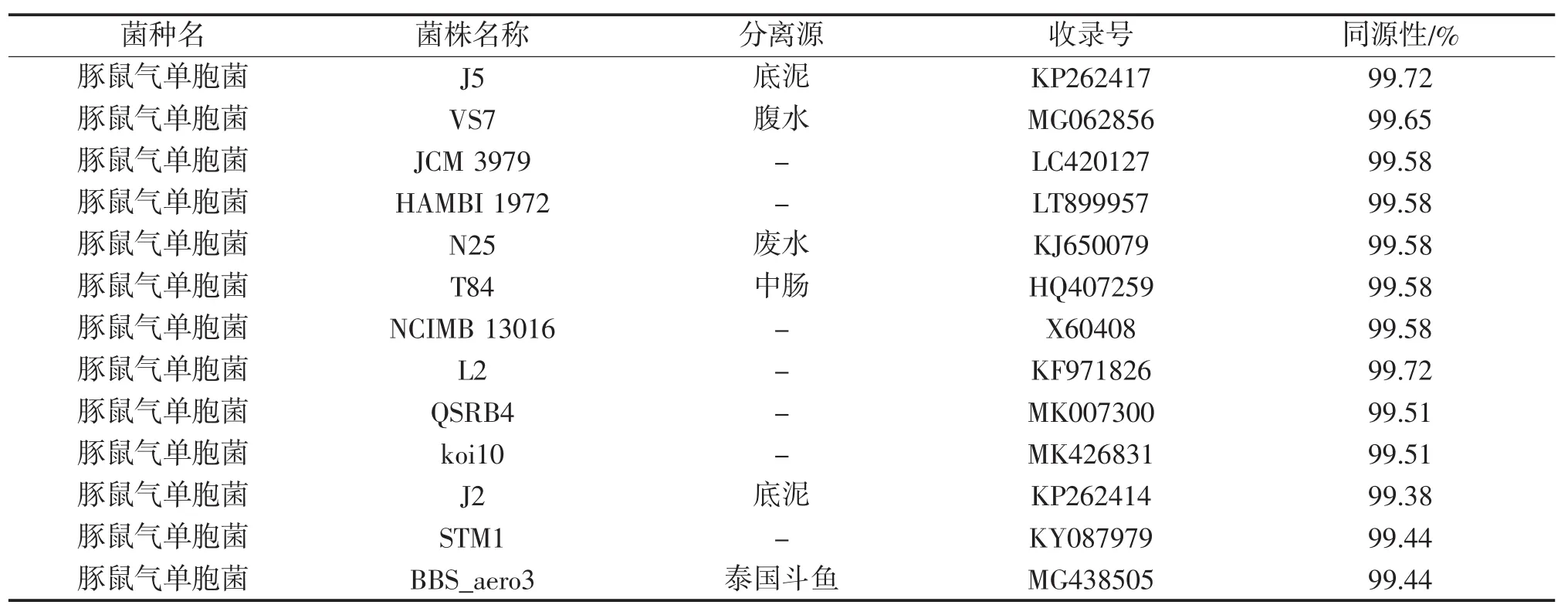
表3 菌株QPX1的16 S rRNA基因序列与GenBank中已知豚鼠气单胞菌菌株16 S rRNA基因序列同源性
2.3 病原菌的药敏特性
菌株QPX1对14种抗生素的敏感试验结果(表4)表明,菌株QPX1对阿莫西林、多粘菌素B、磺胺异噁唑、复方新诺明、卡那霉素、吡哌酸、新霉素、庆大霉素、氟苯尼考、链霉素、恩诺沙星、诺氟沙星、奈替米星等13种抗生素高度敏感,对四环素耐药。
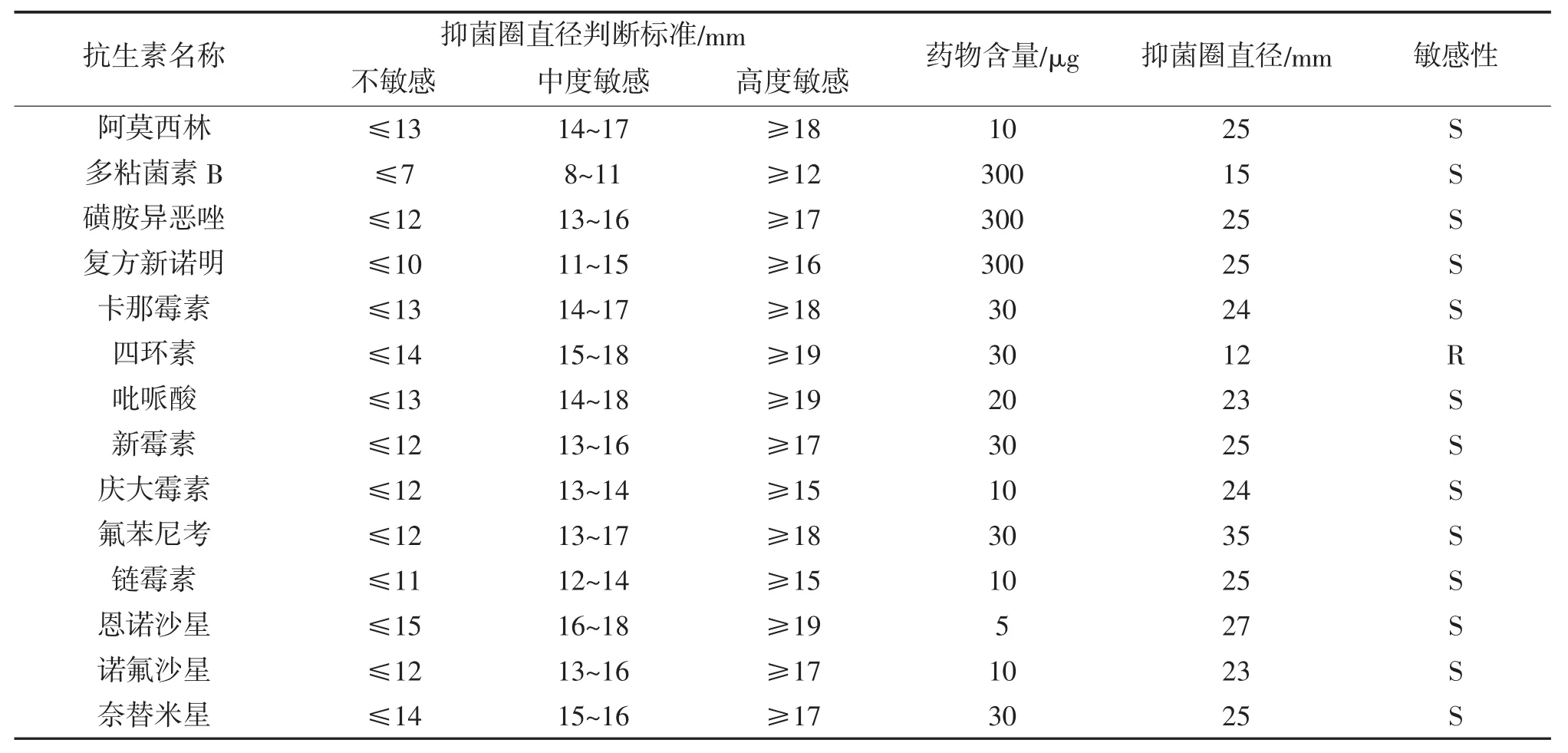
表4 菌株QPX1的药敏特性
3 讨论与分析
3.1 豚鼠气单胞菌对水产养殖的危害性
豚鼠气单胞菌是水产养殖重要病原菌,具有广泛的致病性,已经引起南方鲇(Silurus meriordinalis)[9]、日本鳗鲡(Anguilla japonica)[10]、克氏原螯虾(Procambarus clarkii)[6]、花 鲈(Lateolabrax japonicus)[11]、草鱼(Ctenopharyngodon idellus)[12]、匙吻鲟(Polyodon spathula)[13]等多种水产养殖动物发病死亡,对水产养殖具有极强的危害性。然而,从现有文献资料来看,关于豚鼠气单胞菌对南美白对虾致病的详细报道较少,仅李梅等[5]曾报道豚鼠气单胞菌引起广东养殖南美白对虾发生以“足须发红、肝胰腺肿大”为典型病症的疾病,而豚鼠气单胞菌引起南美白对虾黄腿病的研究尚未报道。该试验对南美白对虾黄腿病病原菌豚鼠气单胞菌进行了分离鉴定,筛选了潜在防控药物,研究结果可作为南美白对虾黄腿病的药物防控提供依据。
3.2 豚鼠气单胞菌的致病机理分析
一般来说,病原菌的致病性与其毒力因子的产生密切相关。据报道[14-15],豚鼠气单胞菌能够产生多种毒力因子。阳涛等[14]实验发现,南方鲇源豚鼠气单胞菌胞外产物具有蛋白酶、脂酶、明胶酶和脲酶活性,具有很强的溶血毒性和细胞毒性;熊静等[15]研究表明,鳗源豚鼠气单胞菌胞外产物具有溶血、酪蛋白酶和卵磷脂酶活性。该试验证实豚鼠气单胞菌对南美白对虾具有致病性,可能与这些毒力因子的产生有关。至于豚鼠气单胞菌引起南美白对虾黄腿病的发病机理,目前尚未确定,可能是由于豚鼠气单胞菌感染南美白对虾后产生毒力因子,引起肌肉等组织出现病变和坏死,导致病虾黄色素病理性沉积,表现出附肢变黄。当然,除了豚鼠气单胞菌的自身因素外,该病的发生与池底腐败有机质累积造成的病原菌滋生、养殖户投喂饲料不当也有较大的关系。
3.3 豚鼠气单胞菌的药敏特性分析
不同来源的豚鼠气单胞菌常常具有药敏特性的差异性。何永龙等[12]从草鱼体内分离的豚鼠气单胞菌菌株JS对恩诺沙星、复方新诺明、氟苯尼考等抗生素高度敏感,对多粘菌素B中度敏感,对阿莫西林耐药;韦阳道等[13]从患病匙吻鲟体内分离的豚鼠气单胞菌菌株GXKJDX02对多粘菌素B、氟苯尼考、复方新诺明等抗生素耐药,对恩诺沙星高度敏感。该试验也对豚鼠气单胞菌菌株QPX1的药敏特性进行了分析,发现菌株QPX1对阿莫西林、多粘菌素B、复方新诺明、氟苯尼考、恩诺沙星等抗生素均高度敏感,与韦阳道等[13]分离的鲟源豚鼠气单胞菌对多粘菌素B、氟苯尼考、复方新诺明等抗生素的敏感性有所不同,与何永龙等[12]分离的草鱼源豚鼠气单胞菌对阿莫西林的敏感性不同。此外,菌株QPX1对磺胺异噁唑、复方新诺明、新霉素、氟苯尼考、恩诺沙星等常见水产用抗生素均高度敏感,说明菌株QPX1对常规水产用抗生素并未产生严重的耐药性,为该病的合理用药防控提供了科学依据。然而,值得指出的是,豚鼠气单胞菌的药敏特性会随着时间的推移均会发生变化,因而非常有必要长期监测豚鼠气单胞菌对常规水产药物的敏感性,以避免临床盲目用药。与此同时,养殖者也应该认清抗生素滥用对豚鼠气单胞菌耐药性产生、水产品质量安全和人类健康造成的潜在危害性,在养殖生产过程中要采用药敏试验筛选有效药物进行抗生素的合理应用[16],严格贯彻和落实农业农村部的兽用抗菌药使用减量行动,推进南美白对虾养殖业的绿色发展。
4 结论
上海青浦养殖南美白对虾黄腿病的病原菌是豚鼠气单胞菌,生产上可选用新霉素、氟苯尼考、恩诺沙星等渔用抗生素进行防治。
猜你喜欢
当代水产(2022年6期)2022-06-29
当代水产(2022年1期)2022-04-26
当代水产(2022年3期)2022-04-26
当代水产(2022年3期)2022-04-26
感染、炎症、修复(2021年1期)2021-07-28
现代畜牧科技(2021年4期)2021-07-21
包头医学院学报(2020年6期)2020-10-22
农家致富顾问·上半月(2019年7期)2019-09-10
阅读(低年级)(2019年6期)2019-08-27
读写算·小学低年级(2016年6期)2016-05-14
