
红掌种质资源重要观赏性状遗传多样性分析
2020-02-22 03:19连子豪黄少华陈艳艳牛俊海尹俊梅
热带作物学报 2020年12期
连子豪 黄少华 陈艳艳 牛俊海 尹俊梅

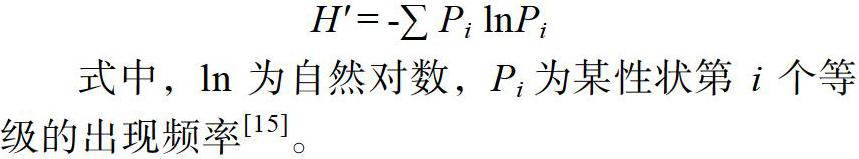
摘 要:为了解红掌种质资源重要观赏性状遗传多样性,以91份红掌种质作为试验材料,对17个重要观赏性状进行鉴定,应用相关性分析、主成分分析和聚类分析等方法分析其遗传多样性。结果表明:供试红掌资源各性状的变异系数为16.27%~66.20%,遗传多样性指数为1.8856~2.0886,说明其具有较高的形态多样性,其中右耳心距变异最大,肉穗花序顶端粗变异最小。相关性分析发现,各观赏性状间存在着较为紧密的正向联系,大部分性状间呈显著或极显著正相关。通过主成分分析,确定的3个主要成分,其累计贡献率为82.991%,由决定植株大小、佛焰苞大小及肉穗花序粗度的指标组成,其中前两个特征对红掌种质多样性影响较大。依据植株大小、佛焰苞大小及肉穗花序粗度进行聚类分析,可将91份红掌种质分为6个类群,各类群特征明显,其中第Ⅵ类群多个性状表现独特,值得特别注意。本研究通过分析红掌种质资源重要观赏性状变异情况,了解其遗传多样性,为红掌遗传改良、种质创新和应用提供理论依据。
关键词:红掌;观赏性状;种质资源;遗传多样性
中图分类号:S682 文献标识码:A
Abstract: In order to understand the genetic diversity of important ornamental traits in the germplasm resources of Anthurium andreanum, 91 germplasm samples were used as the test materials to identify 17 important ornamental traits. The results showed that the variation coefficient of each character of the tested resources ranged from 16.27% to 66.20%, and the genetic diversity index ranged from 1.8856 to 2.0886, indicating that it had higher morphological diversity, among which the right auricular distance variation was the largest and the coarse variation at the top of the inflorescence was the smallest. The correlation analysis showed that there was a close positive correlation among the ornamental traits, and most of the traits showed significant or extremely significant positive correlation. Through principal component analysis, the cumulative contribution rate of the three main components was 82.991%, which was composed of indicators determining plant size, bud size and inflorescence thickness, among which the first two characteristics had a great influence on the diversity of germplasm. According to the size of plants, the size of floriferous bracts and the thickness of inflorescence, the 91 germplasm could be divided into 6 groups, among them, the characteristics of group VI were unique and deserve special attention. This study woul lay a theoretical basis for genetic improvement, germplasm innovation and application of A. andreanum.
Keywords: Anthurium andraeanum; ornamental character; germplasm resources; genetic diversity
DOI: 10.3969/j.issn.1000-2561.2020.12.007
紅掌(Anthurium andraeanum Linden),又名花烛、安祖花等,是天南星科(Araceae)花烛属(Anthurium Schott)多年生常绿花卉。其具有花期长、花型独特、花色丰富等特点,可作为盆栽观赏植物或切花植物。
近些年来,红掌育种技术得到了长足发展和广泛应用,出现了红掌倍性育种[1-3]和分子育种[4-7]等研究的报道,但目前红掌主要的种质改良和品种选育方式仍以传统杂交育种为主[8]。因此,分析红掌的种质资源遗传多样性,对于筛选红掌优良种质、提高育种效率有着重要意义。国内目前对于红掌遗传多样性的研究已有报道,徐世松等[9]对32个红掌品种进行了遗传多样性分析和聚类分析,聚类将其划分为5大类。易双双等[10]对64个红掌品种进行了遗传多样性分析,发现红掌不同表型性状之间存在一定的关系, 绝大部分性状彼此之间呈极显著相关,聚类将其分为6大组群,并发现红掌大小、形态、颜色和来源是分类的主要依据。尚伟等[11]对50个红掌品种进行了11个主要农艺性状多样性研究,发现叶柄长、佛焰苞长、佛焰苞宽和花朵数的变异系数较大,聚类将其划分为4类,并筛选出2个表现较好的类群。植株大小和株型是红掌的重要观赏特征,不仅是人们选择不同红掌时的重要标准,还是红掌育种中的重要目标特征,然而,有关该方面尚未见专门报道。本研究以91份红掌种质作为试验材料,主要针对17个与植株大小、株型和佛焰苞形态相关的重要观赏性状进行分析,旨在挖掘红掌种质资源遗传多样性,对于红掌株型的遗传改良及其新种质的开发具有重要意义。
1 材料与方法
1.1 材料
利用位于海南省儋州市中国热带农业科学院热带作物品种资源研究所热带花卉资源圃的91份红掌种质作为研究材料(表1),其苗龄为3~5 年,在同一个具有完备设施的遮荫大棚内,将供试红掌盆花以25~35 cm的间距整齐摆放在床架上,保证盆土干湿相间的原则,每5 d浇1次水,每14 d施1次复合液肥。棚内温度24~32 ℃,相对湿度60%~80%,光照强度8 000~12 000 lx。
1.2 方法
每个种质选择5株同一时间栽植、株型一致、生长健壮的植株。在红掌佛焰苞的最佳观赏期,即肉穗花序成熟2/3时[12],以中华人民共和国国家标准《植物新品种特异性、一致性和稳定性测试指南?花烛属》和尹俊梅[13]制定的热带花卉资源描述规范为参照标准,调查了株高、花梗长、叶柄长、佛焰苞长、佛焰苞宽、佛焰苞左脸宽、佛焰苞右脸宽、左耳心距、右耳心距、肉穗花序长、叶长、叶宽、肉穗花序顶端粗、肉穗花序中部粗、肉穗花序下部粗、花梗粗、叶柄粗共17个观赏性状,其中长度用直尺和卷尺测量,粗度用游标卡尺测量。数据测量记录时间为2019年6月至2019年8月。
1.3 数据处理
对调查的观赏性状数据计算最小值、最大值、平均值、标准差、变异系数和多样性指数。采用Excel 2019软件进行遗传多样性分析,取调查数据的平均值分析。为了更直观地反映数量关系,便于分析,对调查的观赏性状进行了分级[14],分级方法为:先计算试验材料的总体平均数()和标准差(SD),然后整体划分为10个等级,从第1级[i <(-2 d)]到第10级[i >(+2 d)],每0.5 d划分为1级,通过每一级的相对频率计算出多样性指数。遗传多样性指数的计算公式为:
式中,ln为自然对数,Pi为某性状第i个等级的出现频率[15]。
运用SPSS 23.0[16]软件对17个性状进行相关性分析、主成分分析及聚类分析,其中聚类方法为Ward法,以欧式距离作为品种间距离。
2 结果与分析
2.1 重要观赏性状的遗传多样性分析
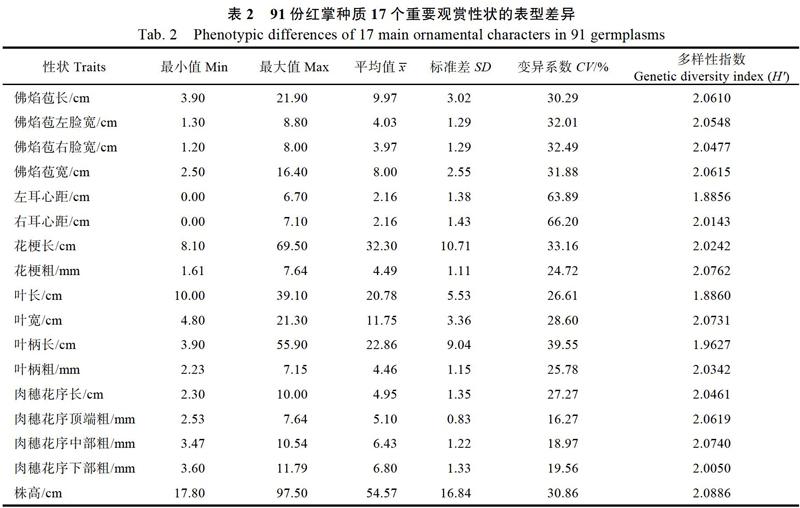
通过对91份红掌种质17个观赏性状的调查,结果表明(表2):供试红掌的17个性状变异系数范围为16.27%~66.20%,其中,右耳心距与左耳心距的变异系数大于60%,叶柄长、花梗长、佛焰苞右脸宽、佛焰苞左脸宽、佛焰苞宽、株高与佛焰苞长的变异系数大于30%,肉穗花序下部粗、肉穗花序中部粗和肉穗花序顶部粗的变异系数小于20%,右耳心距的变异系数最大,肉穗花序顶端粗的变异系数最小。
从表2可见,17个观赏性状的多样性指数范围为1.8856~2.0886,其中株高的多样性指数最大为2.0886,左耳心距的多样性指数数最小为1.8856。
2.2 重要观赏性状间相关性分析
相关性分析结果显示(表3),供试红掌资源的重要观赏性状间存在着较为紧密的关系,且绝大多数为正相关。其中株高与花梗长、叶柄长、叶长、叶宽呈极显著正相关;佛焰苞长与佛焰苞左脸宽、佛焰苞右脸宽、佛焰苞宽、左耳心距、右耳心距、花梗粗呈极显著正相关;肉穗花序顶部粗与肉穗花序中部粗、肉穗花序下部粗、花梗粗呈极显著正相关;花梗粗与叶柄粗呈极显著正相关;花梗长与花梗粗呈极显著正相关;叶柄长与叶柄粗呈极显著正相关。
2.3 红掌观赏性状主成分分析
主成分分析结果显示(表4),17个测量指标中共筛选出了3个主成分,其累计贡献率为82.991%。第1主成分的特征值为5.470,贡献率为32.179%,主要由株高、花梗长、叶柄长、叶长、叶宽、叶柄粗决定,说明第1主成分是由决定植株大小的指标组成。第2主成分的特征值为5.468,贡献率为32.167%,主要由佛焰苞长、佛焰苞左脸宽、佛焰苞右脸宽、佛焰苞宽、左耳心距和右耳心距决定,说明第2主成分是由决定佛焰苞大小的指标所组成。第3主成分的特征值为3.170,贡献率为18.645%,主要由肉穗花序顶部粗、肉穗花序中部粗、肉穗花序下部粗决定,说明第3主成分是由红掌肉穗花序粗度决定的。
2.4 红掌观赏性状聚类分析
聚类分析的结果表明(图1),当欧氏距离为12.5时,可将91份红掌种质分为6大类群。第Ⅰ类群包含42个品种,其特点是:植株整体较小,株型紧凑,适合做盆花;第Ⅱ类群包含11个品种,但与第Ⅰ类群存在较明显区别,其主要特点是:植株较小,但有更具特色的佛焰苞或肉穗花序,在小株型的条件下具有更大的佛焰苞,更粗的肉穗花序或更大的佛焰苞长宽比;第Ⅲ类群包含7个品种,主要特点是:株高较高,花梗、叶柄较长,具有较大的佛焰苞;第Ⅳ类群包含‘甜梦和‘樱桃红2个品种,该类群主要特点是:株高较高,但花梗和叶柄相对较短,佛焰苞与叶片较小;第Ⅴ类群包含28个品种,植株大小介于大型切花和小型盆花红掌之间,其主要特点是:植株大小中等,佛焰苞大小中等,大部分可作为盆花栽培观赏,部分品种可作为切花红掌;第Ⅵ类群中只有1个品种‘Bana 1,该品种花梗短于叶柄,佛焰苞生长常矮于叶片,佛焰苞较大,花梗、叶柄粗壮。
3 讨论
种质资源遗传多样性的研究是育种工作的基础[17]。本研究调查了91份具有代表性的不同优良观赏性状的红掌种质,有针对性地对影响红掌植株大小和株型的17个重要的观赏性状进行遗传多样性分析,结果发现,17个性状的变异系数范围为16.27~66.60,其中右耳心距变异最大;遗传多样性指数范围为1.8856~2.0886。一般情况下变异系数大于10%表示样本间差异大[10],本研究中所有性狀的变异系数均大于10%,表明供试红掌资源的重要观赏性状多样性水平较高,具有较强的改良潜力,有利于后续红掌育种中特异种质材料的筛选工作。
相关性分析结果表明,供试红掌资源17个性状之间绝大部分呈显著或极显著正相关。结合主成分分析结果,筛选出了3个主成分,累计贡献率为82.991%,其反映了17个性状的特征,分别为植株大小、佛焰苞大小和肉穗花序粗度,正是这3个特征的影响造成了红掌种质的多样性,其中第1主成分与第2主成分特征值分别为5.470和5.468,说明株高、花梗长、叶柄长、叶长、叶宽、叶柄粗、佛焰苞长、佛焰苞左脸宽、佛焰苞右脸宽、佛焰苞宽、左耳心距和右耳心距等是新品种选育过程中需要重点考虑的性状,这可以帮助育种者减少选择亲本时考虑的性状数量,简化育种工作。
聚类以植株大小、佛焰苞大小和肉穗花序粗度等为主要依据,将91份红掌种质分为6大类群,供试红掌种质遗传分化明显。第Ⅰ类群:佛焰苞较小的小株型盆花,性状表现较好;第Ⅱ类群:佛焰苞大小中等的小株型盆花,性状表现一般;第Ⅲ类群:佛焰苞较大的大型切花红掌,性状表现较好;第Ⅳ类群性状表现较差;第Ⅴ类群:植株和佛焰苞大小中等可用作盆花或切花,性状表现较好;第Ⅵ类群:为单独聚为一类的‘Bana 1,虽然其肉穗花序矮于叶片,但具有诸多其他红掌品种不具备的特点,如粗壮笔直的花梗,宽大的叶片等,值得特别注意。聚类结果为今后红掌植株大小,株型和佛焰苞及叶片的相对位置等的遗传改良和优良品种的筛选培育提供依据。在育种过程中,可根据育种目标,选择不同的类群作为亲
本。聚类结果与前人研究存在差异,徐世松等[9]对32个红掌品种进行聚类后将其分为5大类;尚伟等[11]通过对50个红掌品种进行聚类后将其分为4大类。分析可能造成差异的原因:(1)选择的种质、数量和苗龄不同;(2)试验环境条件不同,上述两方面因素都会导致聚类结果的不同。
隨着科研技术的不断发展,近年来出现了许多将调查性状所得数据与分子标记手段结合的研究,在花卉领域也得到了发展,如玫瑰、牡丹、睡莲、蝴蝶兰等已见有相关研究。下一步将结合分子标记技术,在分子水平分析红掌种质资源遗传多样性,希望能够更加准确地了解材料,深入发掘红掌种质遗传的调控因子和内在机制,更好地提高育种效率。
参考文献
杜宝贵, 黄丽娟, 张志胜, 等. 红掌花药培养[J]. 生物技术通报, 2009(S1): 189-195.
张志胜, 黎扬辉, 姜 蕾, 等. 红掌四倍体的离体诱导及其鉴定[J]. 园艺学报, 2007(3): 729-734.
田丹青, 潘晓韵, 葛亚英, 等. 秋水仙素离体诱导红掌四倍体试验[J]. 浙江农业科学, 2013(9): 1125-1127.
李小军. 农杆菌介导NPR1基因转化红掌的研究[D]. 上海: 上海师范大学, 2005.
陈艳梅. 红掌高密度SNP遗传连锁图谱构建及疫病抗性佛焰苞花色性状QTL定位[D]. 海口: 海南大学, 2018.
Wang J Y, Chuang K C. Development of novel microsatellite markers for effective applications in Anthurium cultivar identification[J]. Euphytica, 2013, 189(3): 421-431.
马广莹, 史小华, 朱开元, 等. 红掌AaMYB1基因的克隆、表达及异源转化研究[J]. 核农学报, 2017, 31(9): 1708- 1716.
牛俊海, 黄少华, 冷青云, 等. 分子标记技术在红掌研究中的应用与展望[J]. 分子植物育种, 2015, 13(6): 1424- 1432.
徐世松, 王呈丹, 黄素荣, 等. 红掌种质资源形态学标记与遗传多样性分析[J]. 热带作物学报, 2014, 35(10): 1890-1896.
易双双, 杨光穗, 尹俊梅, 等. 红掌主要观赏性状遗传多样性分析[J]. 热带作物学报, 2017, 38(12): 2206-2214.
尚 伟, 陈昌铭, 林发壮, 等. 红掌主要农艺性状多样性分析[J]. 中国农学通报, 2019, 35(6): 129-133.
陈艳艳. 红掌主要观赏性状的QTL[D]. 海口: 海南大学, 2019.
尹俊梅. 热带花卉种质资源描述规范[M]. 北京: 中国农业出版社, 2005: 1-12.
Shannon C E, Weaver W. The mathematical theory of communication[J]. The University of Illinois Urbana, 1949: 3224.
孔繁玲. 植物数量遗传学[M]. 北京: 中国农业大学出版社, 2006.
郝黎仁, 樊元郝, 哲 欧, 等. SPSS实用统计分析[M]. 北京: 中国农业水利水电出版社, 2002: 280-285.
王 娟, 李荫藩, 梁秀芝, 等. 北方主栽燕麦品种种质资源形态多样性分析[J]. 作物杂志, 2017(4): 27-32.
猜你喜欢
福建农业科技(2022年1期)2022-03-13
花火·慧阅读(2021年8期)2021-09-04
青少年日记·小学生版(2017年5期)2017-07-07
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
长江蔬菜·学术版(2016年12期)2017-01-12
吉林农业·下半月(2016年11期)2017-01-09
