
3个三角梅品种的光响应及光合特性
2020-02-22 05:41罗素梅陈鸿宾赖金莉牛王翠钟翠玉肖祖财陈碧露陈旭刘小平
江苏农业科学 2020年23期
罗素梅 陈鸿宾 赖金莉 牛王翠 钟翠玉 肖祖财 陈碧露 陈旭 刘小平


摘要:测定3个三角梅品种的光合参数及其变化规律,并对4个模型的光响应曲线进行拟合效果比较。结果发现,直角双曲线修正模型拟合效果最好;在对3个三角梅品种的光合特性研究中,安格斯对强光、弱光的适应能力及节水能力最强,即耐旱性是最强的,而斑叶红衣女王光合能力是最弱的,推测可能与其斑叶光合色素含量较低的原因有关;3个三角梅品种中,安格斯的光合逆境适应能力最强,在赣南地区具有较高的推广应用价值。
关键词:赣南地区;三角梅;光响应;光合特性;直角双曲线修正模型
中图分类号: S685.160.1 文献标志码: A 文章编号:1002-1302(2020)23-0124-05
三角梅别称叶子花、宝巾花、勒杜鹃,是紫茉莉科(Nyctaginaceae)三角梅属(Bougainvillea)的一类具有极高观赏价值的常绿木本花卉[1],其花色多样,全世界有300多种栽培品种[2],在我国广泛应用于南方城市的园林绿化[3-4]。植物的光合作用可反映植物对光的适应能力和生物量的积累,而光合-光响应曲线是研究植物光合作用的主要方法之一[5-6]。目前,普遍采用的擬合光合-光响应曲线的模型有非直角双曲线模型、直角双曲线模型、直角双曲线修正模型和指数模型[7-8],根据模型拟合出来的光响应曲线可以得出植物的初始量子效率、最大净光合速率、光饱和点、光补偿点及暗呼吸速率,通过这些参数值可以了解植物的光合作用能力[9]。目前,已有部分学者对三角梅的光合特性进行了研究,但主要集中在三角梅光合日变化以及某种胁迫处理下的光合特性研究[10-13],鲜有关于三角梅不同品种光适应性和光合特性等方面的研究,而对三角梅光合-光响应曲线模型的研究还未见报道。本研究以赣南地区长势较好的3个三角梅品种为试验材料,利用Li-6400便携式光合作用测定系统测定其光合作用,比较了非直角双曲线模型、直角双曲线模型、直角双曲线修正模型和指数模型4个不同的光响应曲线拟合模型的拟合效果,并对不同品种三角梅的光合气体交换参数进行分析,筛选出适合三角梅光响应曲线的最佳拟合模型,阐明三角梅的光合生理特性,旨在为筛选出适合赣南地区生长的高光效优良三角梅品种并推广应用提供理论基础。
1 材料与方法
1.1 试验材料
试验材料为三角梅盆栽苗,所选品种为马尼拉小姐(Bougainvillea×buttiana ‘Miss Manila)、斑叶红衣女王(Bougainvillea×buttiana ‘Scarlet Queen Variegated)、伊丽莎白安格斯(Bougainvillea glabra ‘Elizabeth Angus,以下简称安格斯)。试验地位于江西省赣州市花卉研究所苗圃(114°39′53″ E、25°49′12″ N)内,四季常青,年平均气温19.3 ℃,冬无严寒,夏无酷暑,雨量充沛,属中亚热带季风湿润气候,12月平均温度8.8 ℃,7月平均温度28.6 ℃,无霜期 286 d,年平均降水量1 443.2 mm,年均日照时数 1 856.6 h。试验品种于2017年年底从广州市引进,并于温室苗圃内种植,正常供水缓苗并定苗,正常生长1年后开始试验,于2019年7月上旬进行光合作用测定。每个品种挑选3盆长势旺盛的植株,测定的样叶为每盆植株顶部往下第3~5张成熟的功能叶片。
1.2 试验方法
1.2.1 光合-光响应的测定 用 Li-6400便携式光合作用测定系统(Li-cor 6400,美国)测定光合-光响应曲线。试验于晴朗天气的08:30—11:00进行,先将待测植株在无遮阴的露地上诱导30 min,再利用Li-6400-02B LED红蓝光源提供不同强度的光合有效辐射(photosynthetically active radiation,PAR),光合有效辐射设定为0、20、50、75、100、150、200、500、800、1 000、1 200、1 500 μmol/(m2·s),测定时用CO2钢瓶将参比室的CO2浓度稳定在(400.0±0.5)μmol/mol,气体流速Flow值设为(500.0±0.5)mmol/s,设置每次改变光强后最短稳定时间为180 s,最长稳定时间为300 s。测定净光合速率[net photosynthetic rate,Pn,μmol/(m2·s)]、气孔导度(stomatal conductance,Gs,μmol/mol)、胞间CO2浓度(intercellular CO2 concentration,Ci,μmol/mol)、蒸腾速率[transpiration rate,Tr,μmol/(m2·s)],并计算植物水分利用效率(water use efficiency,WUE),计算公式为WUE=Pn/Tr。
1.2.2 拟合模型比较 分别利用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型拟合光响应曲线,计算每个模型初始量子效率α(initial slope)、最大净光合速率[maximum net photosynthetic rate,Pmax,μmol/(m2·s)]、光饱和点[light saturation point,Isat,μmol/(m2·s)]、光补偿点[light compensation point,Ic,μmol/(m2·s)]、暗呼吸速率[dark respiration rate,Rd,μmol/(m2·s)]、决定系数(determination coefficient,R2),并与实测值对比。拟合所用模型[7]的表达式如下:
1.3 数据处理
利用Excel 2007对数据进行整理,SPSS 20.0进行分析,Origin 8.5进行作图。
2 结果与分析
2.1 拟合模型的比较
分别运用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型对马尼拉小姐、斑叶红衣女王和安格斯的光响应曲线参数进行拟合。由表1可以看出,利用直角双曲线拟合出来的马尼拉小姐和安格斯的光补偿点分别为51.553 4、45.944 5 μmol/(m2·s),利用指数方程拟合出来的分别为64.976 2、42.917 7 μmol/(m2·s),远大于两者的实测光补偿点30.0、19.0 μmol/(m2·s)。利用直角双曲线拟合的马尼拉小姐和安格斯的暗呼吸速率分别为4.280、2.357 8 μmol/(m2·s),远大于实测值1.0、0.8 μmol/(m2·s)。利用指数方程拟合出来的斑叶红衣女王的光补偿点为 298.884 6 μmol/(m2·s),远远大于实测值 65.0 μmol/(m2·s);利用直角双曲线拟合的斑叶红衣女王的暗呼吸速率为5.123 5 μmol/(m2·s),远大于实测值0.9 μmol/(m2·s);直角双曲线拟合出来的R2在这几个模型中是最低的。直角双曲线、非直角双曲线和指数方程均不能拟合出光饱和点,只有直角双曲线修正模型可以同时拟合出三角梅的光饱和点和光补償点, 且直角双曲线修正模型拟合出来的各光合参数与实测估计值最接近,此外,就 R2来看, 直角双曲线修正模型的拟合效果最好,因此直角双曲线修正模型是三角梅光响应曲线拟合的最佳模型。
2.2 光响应曲线的拟合
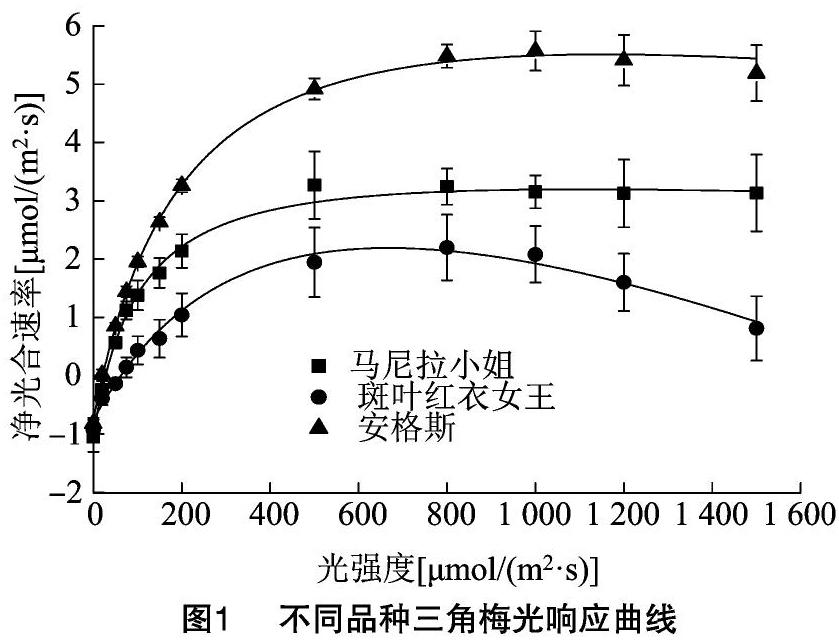
光响应曲线反映植物的光合速率随光强度的变化规律。如图1所示,在一定的光强度范围内,3个三角梅品种的净光合速率随着光强度的增加而升高,当超过光饱和点后,净光合速率随着光强度的增大而降低[14]。其中,在光强度为0~200 μmol/(m2·s)范围内增强时,马尼拉小姐、安格斯和斑叶红衣女王存在一个快速光响应阶段,即净光合速率随着光强度的增大呈线性增长趋势;在光强度为200~800 μmol/(m2·s)范围内增强时,净光合速率随着光强的增大增长幅度逐渐变小;光强度在800~1 500 μmol/(m2·s) 范围内增强时,马尼拉小姐净光合速率随着光强度的增加基本保持不变,安格斯的净光合速率呈降低的趋势,斑叶红衣女王的净光合速率则随着光强度的增加开始下降。由图1可以看出,马尼拉小姐不存在光抑制现象,安格斯在光强度大于1 200 μmol/(m2·s)时存在光抑制现象,斑叶红衣女王则在光强度大于800 μmol/(m2·s)时存在较严重的光抑制现象,高光强度抑制光合作用,导致净光合速率下降,这符合植物正常的生理生长状态。
2.3 气体交换参数随光强度的变化
由图2-A和图2-C可以看出,3个三角梅品种的气孔导度随光强度的变化趋势和蒸腾速率随光强度的变化趋势一致,即安格斯的气孔导度和蒸腾速率随着光强度的增加呈上升趋势,在光强度在0~200 μmol/(m2·s)范围内增长时快速上升,随后上升幅度逐渐变小。马尼拉小姐的气孔导度和蒸腾速率整体上随光强度增大而增加,在光强度为 1 200 μmol/(m2·s) 时存在最大值,之后便下降;斑叶红衣女王的气孔导度和蒸腾速率随光强度的增大先上升后下降,峰值出现在光强度为 75 μmol/(m2·s) 时。
由图2-B可以看出,3个三角梅品种的胞间CO2浓度在光强度在0~200 μmol/(m2·s)范围内增长时呈现急速下降的趋势,之后,斑叶红衣女王的胞间CO2浓度呈现上升的趋势,马尼拉小姐和安格斯也呈上升的趋势,但是上升幅度较小,且处于较平稳的状态。由图2-D可以看出,3个品种的水分利用效率随光强度的变化和胞间CO2浓度随光强度的变化呈现相反的趋势,3个三角梅品种的水分利用效率随着光强度的增加呈先上升后下降的趋势,其中,马尼拉小姐和安格斯在光强度为 150 μmol/(m2·s) 时有最大值,之后随着光强度的增大缓慢下降,而斑叶红衣女王在光强度为 100 μmol/(m2·s) 时有最大值,之后随着光强度的增大急速下降,且在光强度大于 1 000 μmol/(m2·s) 时水分利用效率变为负值。
3 讨论与结论
不同的模型拟合出来的光合参数存在一定的差异,不同的植物应根据拟合结果确定最适合的拟合模型。本研究结果表明,在直角双曲线模型、非直角双曲线模型、直角双曲线修正模型和指数模型4个光响应模型中,直角双曲线修正模型拟合的三角梅的各光合参数的拟合值接近于实测估计值。直角双曲线修正模型下马尼拉小姐、斑叶红衣女王和安格斯的决定系数分别为0.978 5、0.990 7和0.999 4,能够很好地反映这3个三角梅品种的光合速率随光强度的变化趋势。植物的正常生理生长规律是在饱和光强度后净光合速率随光强度增加而降低,也即植物的光响应曲线中是存在一个最大的饱和光强点的,而在这几个模型中,直角双曲线模型、非直角双曲线模型和指数模型不能拟合出光饱和点,只有直角双曲线修正模型可以拟合出三角梅的光饱和强度。叶子飘等发现,直角双曲线新模型能够表现出植物叶片的光抑制现象,且该模型具有普遍性[8];张倚铭等对红花酢浆草的研究发现,直角双曲线修正模型的合理性和精确度最高,光响应曲线的拟合效果最好[15];李永秀等以长江下游冬小麦为研究对象,发现直角双曲线修正模型可以表现植物光抑制的现象,预测效果最好[16];南吉斌等研究发现,直角双曲线修正模型是最佳的光合作用模型[5,17-18]。本研究与前人相似的研究都表明,直角双曲线修正模型对不同植物的光响应曲线的拟合确实具有一定的普遍性,直角双曲线修正模型也是三角梅的最适光响应拟合模型。
通过测定植物的光响应曲线可以探究植物的光合能力,植物光合能力可以通过计算出来的光合参数来衡量。光饱和点能反映不同植物对强光的适应能力[19-20],而表观量子效率和光补偿点反映了植物对弱光的利用情况,表观量子效率越大,光补偿点越低,说明植物利用弱光的能力越强[5]。本研究中,安格斯的Pmax为5.484 4 μmol/(m2·s),光饱和点为977.878 9 μmol/(m2·s),光补偿点为 21.265 7 μmol/(m2·s),表光量子效率最高为 0.037 6,说明安格斯的光适用范围广,对强光和弱光的适应能力较强,对光能利用率较高,并且具有一定的耐荫性。
本研究结果显示,随着光强度的增加,马尼拉小姐和安格斯的气孔导度明显大于斑叶红衣女王,安格斯的最大,说明安格斯气孔限制较小,故其净光合速率也是最大的,馬尼拉小姐其次,斑叶红衣女王最小,因此3个三角梅品种光合能力表现为安格斯 > 马尼拉小姐 > 斑叶红衣女王。斑叶红衣女王光合能力最弱的原因可能是其斑叶叶片内的光合色素含量较低,在一定程度上抑制了光合作用;此外,斑叶红衣女王更容易受到高光强度的抑制,因此在养护管理上要注意长时间的高光强度对其生长的影响。水分利用效率表示单位水量通过蒸腾散失时光合作用所形成的有机物的量[21]。植物的水分利用效率越大,说明植物节水能力越强,耐旱能力和适应能力越强[22]。本研究中,随着光强度的增加,安格斯的水分利用效率一直处于高于马尼拉小姐和斑叶红衣女王的状态,说明安格斯对环境的适应能力最强,其次是马尼拉小姐,最后是斑叶红衣女王,这个结果与光响应曲线的分析结果相一致。
综合可知,直角双曲线修正模型可作为三角梅光合-光响应曲线研究的最佳模型;在对3个三角梅品种光合特性的研究中,安格斯对强光、弱光的适应能力及节水能力最强,即耐旱性是最强的,而斑叶红衣女王光合能力是最弱的,推测可能与其斑叶光合色素含量较低有关;3个三角梅品种中,安格斯的光合逆境适应能力最强,在赣南地区具有较高的推广应用价值。
参考文献:
[1]周 群.三角梅栽培与鉴赏[M]. 北京:金盾出版社,2009.
[2]武晓燕.叶子花属观赏种质资源的遗传多样性分子评价及亲缘关系分析[D]. 厦门:华侨大学,2012.
[3]王晗璐.广西北海市花公园设计与三角梅园林应用[J]. 工程建设与设计,2019,402(4):41-42.
[4]花 老.不同种属三角梅着花特点与园林应用[J]. 中国花卉园艺,2018,426(18):36-38.
[5]南吉斌,杨广环,赵玉文,等. 西藏沙棘属3种植物光合特性及光响应曲线拟合研究[J]. 高原农业,2019,3(3):255-261,295.
[6]刘建锋,杨文娟,史胜青,等. 崖柏与侧柏光合特性和叶绿素荧光参数的比较研究[J]. 西北植物学报,2011,31(10):145-151.
[7]叶子飘,王建林.植物光合-光响应模型的比较分析[J]. 井冈山大学学报(自然科学版),2009,30(2):9-13.
[8]叶子飘,于 强.一个光合作用光响应新模型与传统模型的比较[J]. 沈阳农业大学学报,2007,38(6):771-775.
[9]Sharp R E,Matthews M A,Boyer J S.Kok effect and the quantum yield of photosynthesis:light partially inhibits dark respiration[J]. Plant Physiology,1984,75(1):95-101.
[10]李永红,杨 悦.水分胁迫对叶子花光合特性的影响[J]. 北方园艺,2009(1):191-194.
[11]张学权.2个三角梅品种光合生理特性比较分析[J]. 成都大学学报(自然科学版),2011(3):11-13,29.
[12]唐源江,武小燕.几个三角梅品种光合作用日变化的比较研究[J]. 北方园艺,2011(12):82-85.
[13]施建羽.盐胁迫对三角梅光合特性的影响[J]. 福建热作科技,2012,37(1):5-8.
[14]于 强,叶子飘.光合作用光响应模型的比较[J]. 植物生态学报,2008,32(6):1356-1361.
[15]张倚铭,黎云祥,余茂蕾.红花酢浆草光合模型适用性研究[J]. 生态科学,2018,134(2):20-26.
[16]李永秀,杨再强,张富存.光合作用模型在长江下游冬麦区的适用性研究[J]. 中国农业气象,2011,32(4):588-592.
[17]赵玉文,段少荣,郑 雨.几种光合作用模型下砂生槐光响应和CO2响应曲线的比较[J]. 高原农业,2017,2(2):30,58-64.
[18]李国良,林赵淼,许泳清,等. 不同类型甘薯光合光响应曲线拟合及比较分析[J]. 福建农业学报,2018,33(7):687-690.
[19]高贵宾,钟 浩,吴志庄,等. 不同混生地被竹光合和荧光特征比较[J]. 福建农林大学学报(自然科学版),2016,45(5):515-521.
[20]李 君,娄运生,马 莉,等. 夜间增温和水分管理耦合对水稻叶片光合作用和荧光特性的影响[J]. 江苏农业学报,2019,35(3):506-513.
[21]胡化广,张振铭,吴生才,等. 植物水分利用效率及其机理研究进展[J]. 节水灌溉,2013(3):11-15.
[22]孔文娟. 4个油茶品种的光合特性研究[D]. 兰州:甘肃农业大学,2013.庞一波,刘也楠,吴 江,等. 花前摘心和赤霉素处理对葡萄花序拉长的影响[J]. 江苏农业科学,2020,48(23):129-133.
猜你喜欢
作文小学中年级(2023年1期)2023-02-12
作文新天地(小学版)(2021年8期)2021-08-24
风流一代·青春(2019年6期)2019-06-19
快乐语文(2018年33期)2018-11-29
新教育(2018年8期)2018-08-29
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
椰城(2016年5期)2016-08-04
