
抑郁症大鼠海马神经元自噬与PTEN的表达
2020-02-19 05:20赵俊华王海涛艾莉波郭艳华徐爱军
中风与神经疾病杂志 2020年1期
赵俊华, 王海涛, 刘 浩, 艾莉波, 郭艳华, 徐爱军
目前抑郁症(depression)是第二大导致全球疾病负担的疾病[1],但发病机制尚未明确。海马体主要负责记忆和情感控制,抑郁症会造成海马损伤体积缩小。研究发现抑郁症模型大鼠海马神经元存在显著自噬[2,3];抑郁症模型动物突触明显损伤并伴有数量减少[4];抑郁症海马神经元突触可塑性显著改变[5],这些改变可能是造成其海马神经元损伤及抑郁症发病的重要因素。第10号染色体上缺失的磷酸酶和张力蛋白同源物(PTEN)的脂质磷酸酶活性[6~8],主要可使三磷酸肌醇(PI3P)脱磷酸为二磷酸肌醇(PI2P),从而拮抗磷脂酰肌醇 3 磷酸激酶(PI3K),通过抑制PI3K/ Akt /mTOR 信号通路的作用[9],诱导细胞自噬,为自噬的正调控因子[10]。突触前自噬可促进突触功能和突触的可塑性[11]。而PTEN也参与了成年后突触可塑性过程[12]。PTEN在突触上直接作用的改变可能导致突触功能障碍,从而导致行为和认知后果[13]。研究[14]发现抑制PTEN可以改善正常的突触功能和认知,过表达PTEN的转基因小鼠表现出突触抑制。可以推测抑郁症海马神经元自噬、突触可塑性改变以及PTEN蛋白的表达间可能互有关联,然而尚未见有研究报道。本研究通过建立抑郁症大鼠模型,检测自噬相关因子LC3,Beclin1以及PTEN,观察海马神经元自噬时是否存在PTEN的异常表达并探讨相关性,为理解抑郁症发病机理寻找新的视角。
1 材料与方法
1.1 试剂和仪器 引物、SDS-PAGE凝胶试剂盒为北京庄盟国际生物基因科技有限公司合成;β-actin兔单克隆抗体为ABclonal公司生产;LC3兔多克隆抗体为MBL公司生产;PTEN小鼠单克隆抗体和Beclin1小鼠单克隆抗体为Santa cruz biotechnology公司生产。蛋白电泳仪及化学发光拍摄系统为Bio-RAD公司制造;激光共聚焦显微镜为Leica公司生产;实时荧光定量PCR仪为Roche公司生产。
1.2 动物分组及建立抑郁症模型 取成年健康雄性SD大鼠共50只,体重180~200 g,由华北理工大学实验动物中心提供,动物合格证编号:SCXK(京)2014-0004。适应性饲喂7 d。随机分为2组(模型组和对照组),25只每组。模型组通过慢性不可预见温和性应激(CUMS)[15]方法制备抑郁症大鼠模型。应激包括11种,共持续21 d。每天随机给与1到2种,每种应激不超过3次,同种应激方法不连续出现。对照组大鼠仅每天抓取一次。
1.3 行为学 旷场实验检测对陌生环境的探索欲:记录其主动逃避时间和轨迹。Morris 水迷宫(MWM)检测对寻找空间位置的学习和记忆能力:含隐蔽站台实验和空间探索实验。隐蔽站台实验,记录其游泳轨迹及寻找站台时间,即逃避潜伏期(escape latency,EL)。隐蔽站台实验24 h后进行空间探索实验,撤除站台,记录游泳轨迹、穿越原站台所在位置次数。悬尾不动实验检测抑郁状态程度:对不动时间进行统计。糖水偏好实验评价动物快感缺失的程度:快感缺失的出现是抑郁症动物模型造模成功的标志。记录大鼠的糖水和纯水消耗以及总液体消耗。糖水偏好指数% =糖水消耗量/(糖水消耗量+纯水消耗量)×100%。
1.4 免疫组织荧光 按不同时间点(0.5 d、1 d、3 d、7 d、14 d)分别将2组大鼠10%水合氯醛麻醉后,4 ℃预冷4%多聚甲醛经心脏主动脉灌注固定,断头后冰上快速取脑。4%多聚甲醛4 ℃固定24 h后15%、20%、30%蔗糖梯度沉糖脱水。OCT包埋,-80 ℃冰冻,25 μm冰冻切片。10%胎牛血清室温封闭45 min,一抗(LC3,1∶500;Beclin1,1∶500;PTEN,1∶500)37 ℃湿盒孵育1 h。荧光二抗IgG(1∶100)37 ℃湿盒避光孵育1 h。带DAPI封片剂染核封片。激光共聚焦显微镜观察。
1.5 Western blot 按不同时间点(0.5 d、1 d、3 d、7 d、14 d)分别将2组大鼠麻醉后断头取脑。冰上快速分离双侧海马,-80 ℃冻存,一侧用于提取组织总蛋白,另一侧用于组织总RNA提取。BCA法蛋白定量后按照30 μg蛋白量上样,进行15%聚丙烯酰胺凝胶电泳,PVDF膜电转印,8%脱脂奶粉封闭,一抗(LC3,1∶1000;Beclin1,1:1000;PTEN,1∶1000;β-actin,1∶10000)室温孵育2 h,二抗IgG(1∶5000)室温孵育1 h,ECL法显色。LC3的表达水平以LC3-Ⅱ/LC3-Ⅰ比值表示。目的蛋白的表达水平以目的条带与内参蛋白的光密度比值表示。
1.6 实时荧光定量PCR 取前述冻存的一侧海马组织,提取总RNA,用超微量分光光度计测定RNA浓度和纯度。后参照试剂盒方案进行反转录。实时荧光定量PCR反应条件94 ℃变性15 s,60 ℃退火30 s,72 ℃延伸30 s,共40个循环。
2 结 果
2.1 行为学检测结果 旷场实验:与对照组比较,模型组大鼠总路程、直立次数减少,结果有统计学意义(P<0.01,见图1A、1B、1C)。
水迷宫实验:与对照组相比,模型组大鼠显示显著的逃避潜伏期延长及总路程增加,结果有统计学意义(P<0.01,见图2A、2C、2D);穿台次数显著减少,结果有统计学意义(P<0.01,见图2B、2E)。
穿梭箱实验:与对照组相比,模型组大鼠主动逃避时间增加,结果有统计学意义(P<0.01,见图3A、3B)。
悬尾不动实验:与对照组相比,模型组大鼠悬尾不动时间显著增加,结果有统计学意义(P<0.01,见图4A)。糖水偏好实验:与对照组相比,模型组大鼠糖水消耗比例明显降低,结果有统计学意义(P<0.01,见图4B)。
2.2 免疫组织荧光检测结果 荧光镜下:与PTEN和 Beclin1相结合的是Rhodamine标记的二抗,594nm波长激光下发红光。与LC3相结合的是Fluorescein的二抗,488nm波长激光下发绿光。红色和绿色荧光二者叠加呈黄色,表示该部位存在目的蛋白共定位现象。可见模型组海马神经元PTEN与LC3蛋白呈共定位表达,Beclin1与LC3蛋白也呈共定位表达。与对照组相比,模型组红绿色荧光亮度均较强,同时PTEN蛋白的表达量也增加(见图5)。
2.3 Western blot结果 蛋白质免疫印迹(见图6A):与对照组相比,模型组大鼠海马PTEN、Beclin1与LC3的表达量在相同时相点均增高,结果具有统计学意义(P<0.01,见图6B、6C、6D);与对照组相比,LC3-Ⅱ/LC3-Ⅰ的比值呈随时间递增趋势,自0.5 d时小于1,至14 d时的大于1的翻转,结果有统计学意义(除1 d、3 d外P<0.01,见图6E)。对照组内LC3-Ⅱ/LC3-Ⅰ的值不同时相点两两相比无显著差异(P>0.05,见图6E),模型组组内各时相点LC3-Ⅱ/LC3-Ⅰ的值两两相比(除1 d与3 d间相比无显著差异外)差异显著,均有统计学意义(P<0.05,见图6E)。
2.4 实时荧光定量PCR检测结果 大鼠海马组织自噬相关基因Beclin1、LC3与PTEN的mRNA表达水平与对照组相比,各时相点模型组大鼠海马组织Beclin1、LC3与PTEN的mRNA表达水平均增加,以Beclin1和LC3的mRNA增加更显著,PTEN的mRNA增加次之,组间差异显著,结果具有统计学意义(P<0.01,见图7A、7B、7C)。与对照组相比,模型组内各时相点Beclin1、LC3的mRNA表达水平呈随时间的下降趋势,各时相点间差异显著,差异有统计学意义(P<0.01,见图7A、7B);而模型组内PTEN的mRNA各时相点间表达水平无显著差异(P>0.05,见图7C)。
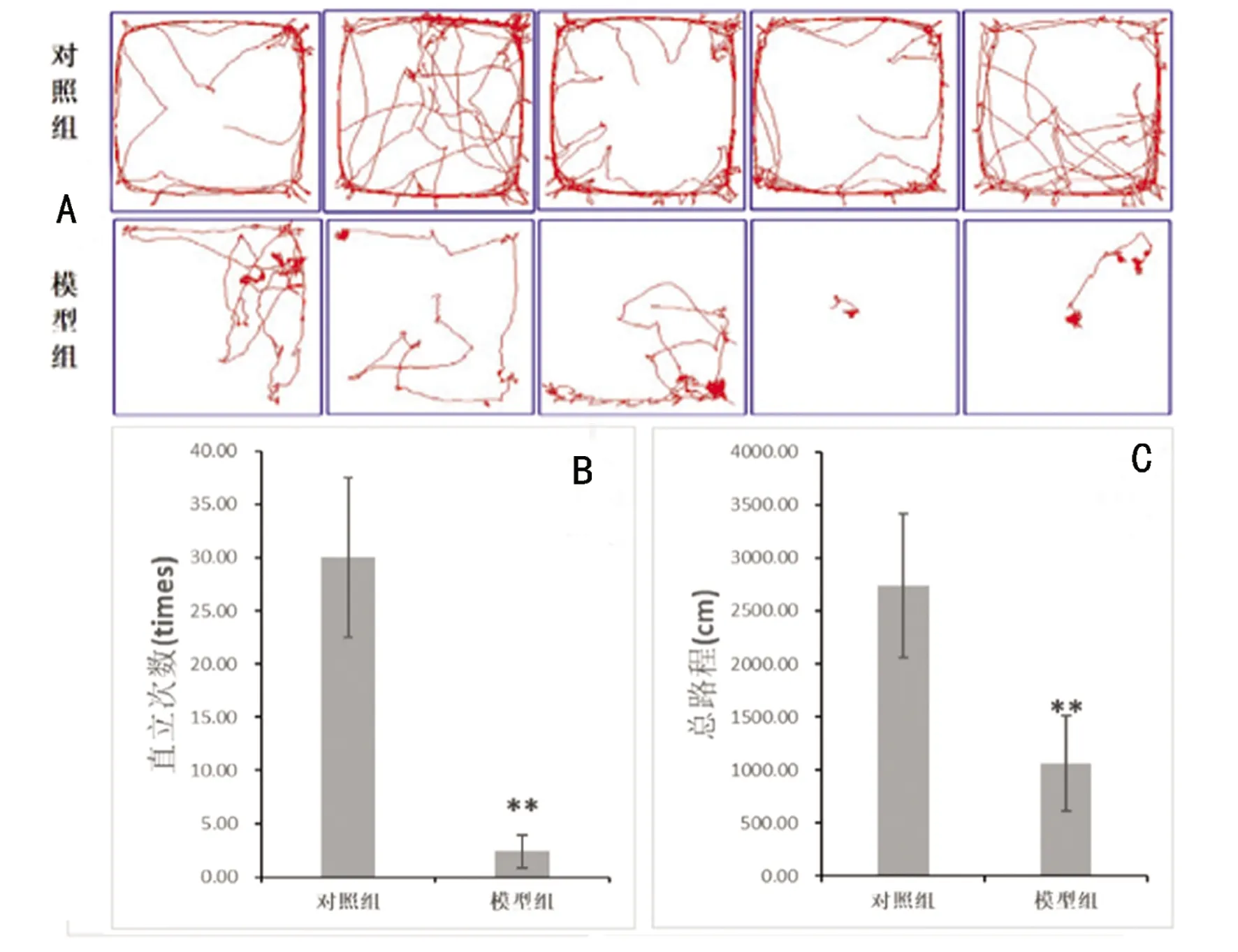
A:旷场实验各组轨迹(n=5);B:旷场实验各组大鼠直立次数比较(n=5),**与对照组相比P<0.01;C:旷场实验各组总路程比较(n=5),**与对照组相比P<0.01
图1 旷场实验(n=5)
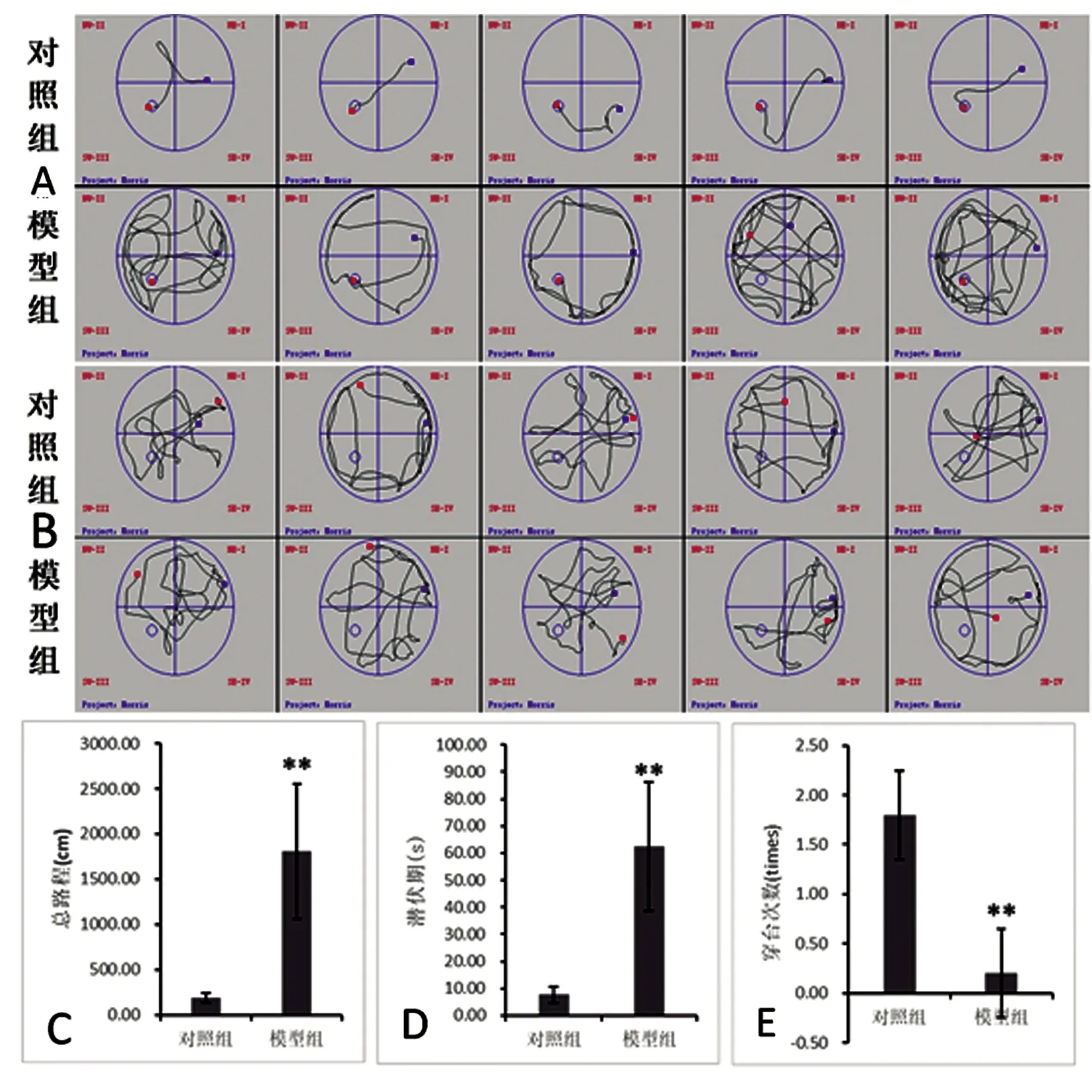
A:MWM实验轨迹图(n=5);B:MWM隐蔽站台实验轨迹图(n=5);C:MWM 实验各组总路程比较(n=5),**与对照组相比P<0.01;D:MWM 实验各组逃避潜伏期比较(n=5),**与对照组相比P<0.01;E:MWM 实验各组穿越原站台次数比较(n=5),**与对照组相比P<0.01
图2 MWM实验(n=5)
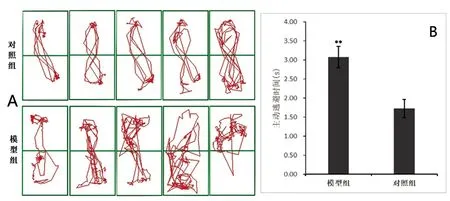
A:穿梭箱实验各组轨迹比较(n=5);B:各组大鼠平均主动逃避时间比较(n=5),**与对照组相比P<0.01
图3 穿梭箱实验(n=5)
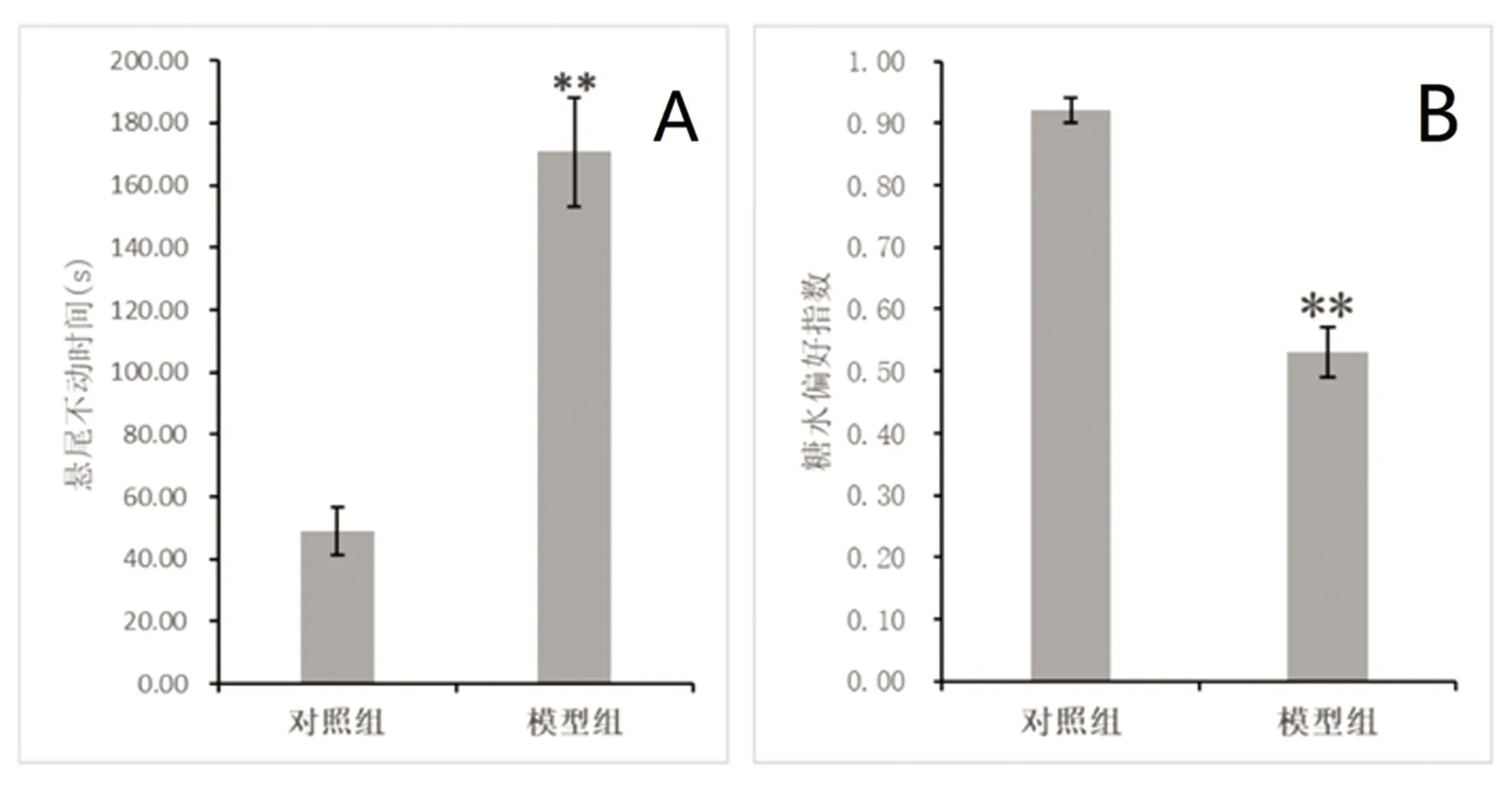
A:悬尾实验中各组大鼠的平均不动时间的比较,**与对照组相比P<0.01;B:糖水偏好实验各组大鼠糖水偏好指数比较,**与对照组相比P<0.01
图4 悬尾实验和糖水偏好实验(n=5)
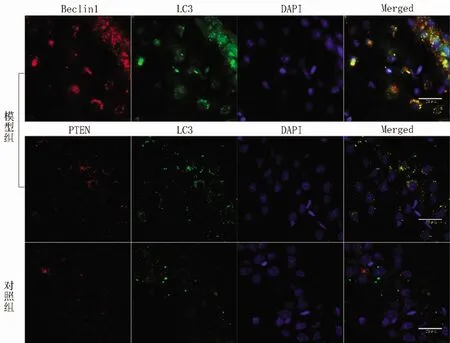
图5 免疫荧光双标检测各组 PTEN 和 Beclin1 分别与LC3共定位表达(n=5)
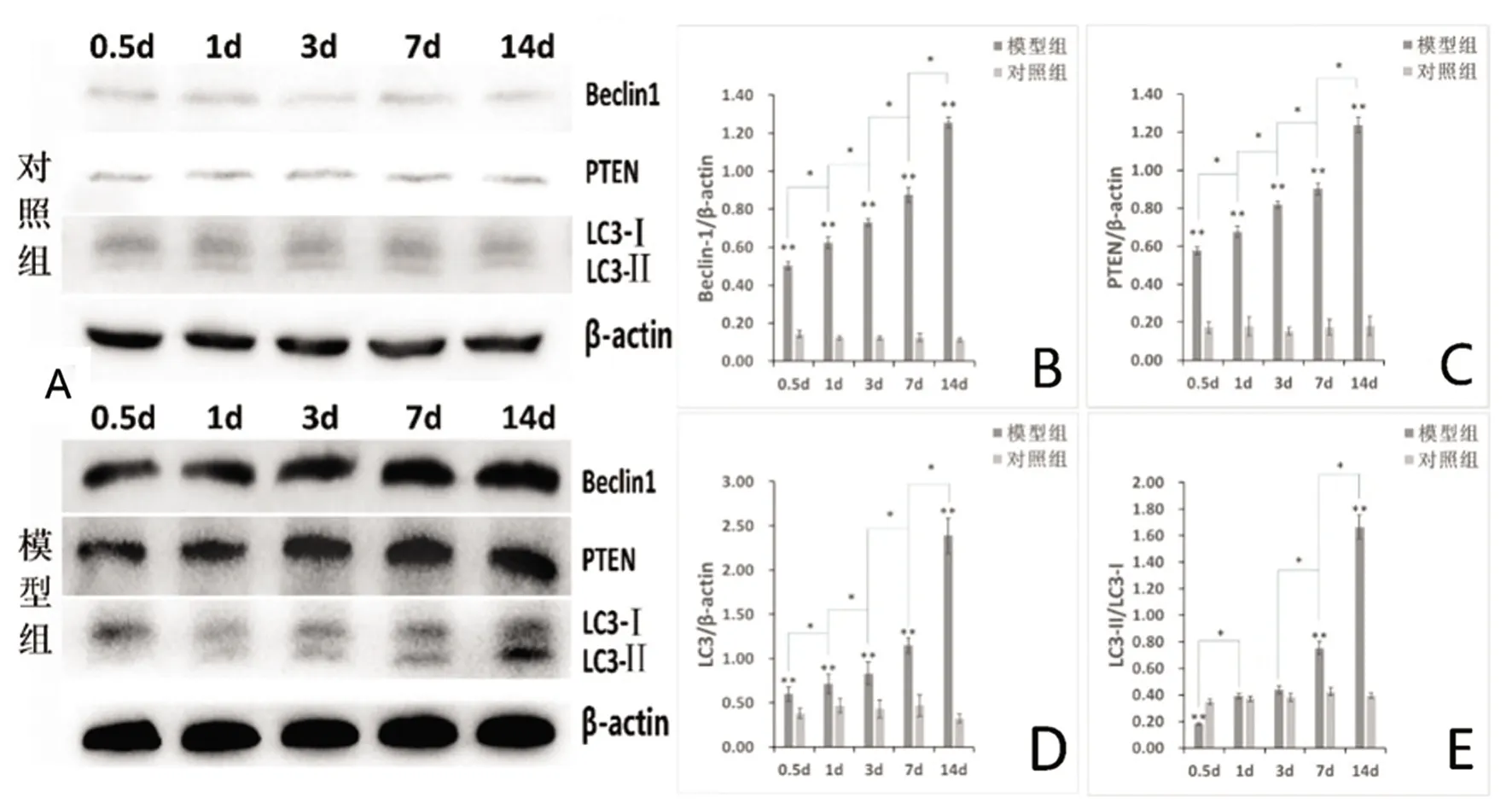
A:Western blot 各组大鼠海马不同时相点各目蛋白的表达;B:模型组各时相点大鼠海马组织Beclin1蛋白表达水平均上升,**与对照组相比P<0.01,*模型组内各相邻前后时相点间相比,Beclin1蛋白表达水平均升高,P<0.01;C:模型组各时相点大鼠海马组织PTEN蛋白表达水平均上升,**与对照组相比P<0.01,*模型组内各相邻前后时相点间相比,PTEN蛋白表达水平均升高,P<0.01;D:模型组各时相点大鼠海马组织LC3蛋白表达水平均上升,**与对照组相比P<0.01,*模型组内各相邻前后时相点间相比,LC3蛋白的表达水平均升高,P<0.01;E:模型组各时相点大鼠海马组织LC3-Ⅱ/LC3-Ⅰ比值呈随时间递增趋势,**与对照组相比P<0.01,*模型组内各相邻前后时相点间相比P<0.01
图6 Western blot各组大鼠海马不同时相点各目蛋白的表达(n=5)

A:模型组各时相点大鼠海马组织LC3的mRNA表达水平均上升,**与对照组相比**P<0.01,*模型组内各相邻前后时相点间相比,LC3的mRNA表达水平降低,P<0.01;B:**与对照组相比P<0.01,模型组各时相点大鼠海马组织Beclin1的mRNA表达均上升,*模型组各相邻前后时相点间相比P<0.01,Beclin1的mRNA表达水平降低;C:**与对照组相比P<0.01,模型组各时相点大鼠海马组织PTEN的mRNA表达水平均上升,*模型组各相邻前后时相点间相比P>0.05,PTEN的mRNA表达无显著差异
图7 LC3、Beclin1及PTEN的mRNA表达水平(n=3)
3 讨 论
本研究采用CUMS应激建立大鼠抑郁症动物模型更加真实的模拟了抑郁症自然发病的过程,更为接近抑郁症发病的真实情况[16]。采用多种行为学方法从多方面进行验证:旷场实验中模型组大鼠对新环境的好奇心缺失;水迷宫实验显示模型组大鼠空间学习记忆能力受损,长期记忆能力下降;穿梭箱实验可见模型组行动轨迹与对照组相比明显杂乱,说明大鼠学习记忆能力较对照组明显下降;悬尾实验中模型组大鼠存在明显的行为绝望状态;糖水偏好实验显示模型组大鼠存在明显的快感缺失。这些结果均支持大鼠抑郁症造模成功可靠。
本研究采用实时荧光定量PCR及蛋白质免疫印迹法分别检测了大鼠海马神经元目的 基因的mRNA及蛋白表达情况。结果显示与对照组相比,模型组大鼠海马神经元两种自噬相关蛋白LC3与Beclin1与其mRNA表达水平均升高,显示其自噬水平增高,提示抑郁症大鼠海马神经元细胞内损伤细胞器及待降解的蛋白质增加,通过增强自噬来清除和纠正。模型组各时相点LC3总蛋白及LC3-Ⅱ蛋白表达呈随时间上升趋势,但实时荧光定量PCR结果中LC3的mRNA的表达呈随时间下降趋势,推测抑郁症大鼠海马神经元LC3的转录环节与翻译环节存在某种未知机制的负反馈。LC3的mRNA表达最高时(0.5 d)LC3总蛋白及LC3-Ⅱ蛋白表达较低,至LC3的mRNA表达最低时(14 d)LC3总蛋白及LC3-Ⅱ蛋白表达反而较高,推测LC3基因在翻译环节存在阻滞,导致了翻译效率降低,阻滞在应激解除后随时间增加而逐渐减轻,LC3总蛋白及LC3-Ⅱ蛋白表达增加。
另外蛋白质免疫印迹显示对照组不同时相点LC3-Ⅱ/LC3-Ⅰ的值相对恒定,且各时相点LC3-Ⅱ蛋白的表达均低于LC3-Ⅰ的蛋白表达。而模型组LC3-Ⅱ蛋白从0.5 d时相点的微量表达显且著低于同时相LC3-Ⅰ蛋白的表达,至14 d时相点的LC3-Ⅱ蛋白表达显著升高并高于同时相LC3-Ⅰ的逆转,推测抑郁症大鼠应激过程中海马神经元可能存在LC3-Ⅰ向LC3-Ⅱ转化环节的阻滞,并可能导致了自噬过程的障碍,解除应激后这种阻滞随时间增加而趋于纠正。另一种自噬相关基因Beclin1的转录及翻译水平随时间变化的趋势亦与LC3的转录及翻译水平变化趋势类同,同样佐证了上述猜想。
实时荧光定量PCR及蛋白质免疫印迹结果显示,抑郁症模型组大鼠海马神经元PTEN基因转录和翻译水平与对照组相比均上调。PTEN蛋白表达也呈上升趋势,与Beclin1蛋白表达时间趋势相近。前人研究表明PTEN可对自噬进行调控[17,18],PTEN为细胞自噬的正调控因子[19]。因此推测PTEN蛋白表达上调对抑郁症大鼠海马神经元自噬起到了协同或促进的作用。与对照组相比,模型组各时相点的大鼠海马神经元PTEN的mRNA均上调,但并未出现与PTEN蛋白表达相似的随时间变化趋势,而是呈平稳的状态,推测PTEN基因的翻译环节也存在一定程度的阻滞。PTEN位于通路的上游,对自噬的调节效应可能会经由通路各环节逐级放大,或可解释PTEN的mRNA上调的程度与Beclin1及LC3的mRNA相比较低的现象。这一点也使得PTEN可能成为对自噬调节更为灵敏高效的靶点。
本研究组织免疫荧光双标方法观察自噬相关蛋白(Beclin1及LC3)与PTEN蛋白的表达。结果显示在抑郁症模型组大鼠海马神经元PTEN 和Beclin1分别与LC3存在共定位,推测PTEN蛋白与Beclin1也存在共定位。这些现象也提示PTEN很可能对抑郁症大鼠海马神经元自噬发生与调控起了到重要的作用。
综上所述抑郁症大鼠海马神经元自噬增强现象与PTEN表达上调有关。PTEN可能是抑郁症大鼠海马神经元自噬改变的重要调控因素之一。另外PTEN也可能经由自噬途径造成海马神经元突触的重塑。这些均有待进一步深入研究。
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
初中生学习指导·中考版(2021年2期)2021-09-10
电子产品世界(2021年8期)2021-01-16
今日农业(2020年13期)2020-12-15
初中生世界·七年级(2020年6期)2020-09-03
时代邮刊(2019年16期)2019-07-30
中国疼痛医学杂志(2019年9期)2019-01-04
创新时代(2016年8期)2016-10-21
科技知识动漫(2016年6期)2016-06-24
汽车观察(2009年1期)2009-02-18
