
黄牛木种群生长模型及动态模拟*
2020-02-12 06:57韦彩丽谢正生孔令华何晓慧
林业与环境科学 2020年4期
韦彩丽 谢正生 孔令华 何晓慧
(华南农业大学 林学与风景园林学院,广东 广州 510642)
黄牛木(Cratoxylum cochinchinense)是金丝桃科(Hypericaceae)黄牛木属(Cratoxylum)的灌木或小乔木,是我国南方少数地区的特有野生树种,喜湿润、酸性土壤,常见于丘陵、山地的疏林或灌丛中,生长慢但萌芽力强;黄牛木木材坚硬珍贵,树干黄褐色,花序聚伞状,花朵粉嫩微香,是优良的用材树种、蜜源植物及城市绿化树种[9]。然而,黄牛木野生资源少,天然更新能力差,我国仅广东、广西、云南有少数分布。目前,尚未发现关于黄牛木引种育苗方面的报道,种群生态学方面的研究也鲜见发表,其研究主要集中于成分分析和药用价值方面[10-12]。本研究通过对黄牛种群生长因子的调查研究,分析其树高、冠幅、数量与胸径的相关性,构建种群生长因子相互关系模型,绘制黄牛木种群生长动态三维模拟图,直观地反映种群生长动态规律及环境适应机制,为黄牛木天然林的抚育管理和经营保护提供科学参考,同时为该地区开展森林资源调查、林分生长研究以及林相结构调整等提供理论基础,从而促进更多优良树种在城市林业和景观生态方面的应用和推广。
1 研究材料及方法
1.1 样地设置与外业调查
在林分踏查的基础上,采用典型样地调查,于2017 年在华南珠三角地区布设3 个黄牛木天然林样地,其中,在广州市的茶山公园和白云山风景名胜区分别选取3 900 m2,在台山市的石花山森林公园选取3 200 m2,样地总面积为11 000 m2。调查对样地内所有黄牛木个体的胸径、树高、冠幅、枝下高等特征进行测定,并记录每株黄牛木在样地内的相对坐标,同时记录各样地的经纬度、海拔、坡度、坡向等环境因子。
1.2 数据分析
1.2.1 种群生长因子统计 采用“空间差异代替时间变化的方法”,即胸径大小代替年龄结构的方法[13-14]来统计分析种群生长因子有关数据,探讨其种群数量变化动态。本研究借鉴有关种群大小级划分方法[15-16],把胸径<1 cm 的黄牛木幼树记为1 级(含树高<1.3 m 的黄牛木个体),胸径≥1 cm、每隔2 cm 为一个龄级,将黄牛木划为11 个径阶,第1 径阶:D<1 cm,第2 径阶:1 ≤D<3 cm,第3 径 阶:3 ≤D<5 cm,第4 径 阶:5 ≤D<7 cm,第5 径 阶:7 ≤D<9 cm,第6 径阶:9 ≤D<11 cm,第7 径阶:11 ≤D<13 cm,第8 径阶:13 ≤D<15 cm,第9 径阶:15 ≤D<17 cm,第10 径阶:17 ≤D<19 cm,第11 径阶:19 ≤D<21 cm。参照林木发育阶段的划分标准[17],将不同胸径大小的黄牛木个体划分为5 个不同的龄组,分别为:幼树(第1 径阶,D<1 cm)、小树(第2~3 径阶,1 ≤D<5 cm)、中树(第4~6 径阶,5 ≤D<11 cm)、大树(第7~8 径 阶,11 ≤D<15 cm)、老 树(第9~11 径阶,15 ≤D<21 cm)。
1.2.2 种群生长因子关系模型构建 一般认为,树种的胸径与年龄、树高、冠幅等生长因子之间存在一定的相关性,且为正相关[18]。年龄与胸径的相关性研究报道已有不少,如此前推出的Richards生长曲线y=A(1-e-kx)B已得到各界的认可并频繁使用,谢正生等[20]通过对广州白云山林分生长与功能的研究,总结出降真香等6 种阔叶树种的胸径与年龄方程,其中,降真香胸径(Y)-年龄(x)生长曲线模型为Y=21.346 5(1-e-0.033133x)1.240376(R2=0.994 9);龙汉利等[19]对红椿(Toona ciliata)胸径与年龄的相关性进行拟合,得出方程:D=2.036 6 ln 0.781(R2=0.759 5)。前人的研究均表明年龄与胸径显著正相关,不再赘述。本研究将对树高与胸径、冠幅与胸径、数量与胸径的相关性进行深入探究,主要运用SPSS 软件对调查数据进行回归分析,在曲线估计下将胸径设置为自变量,树高、冠幅、数量设置为因变量,得到黄牛木生长因子之间相互关系的曲线模拟图以及模型参数估计值。通过比较模型参数估计值的相关性显著系数,结合拟合曲线以及树种的生物学特性来选取最佳拟合模型,即系数R2越大,表明两者相关性越显著,拟合程度越高;方差分析显著水平P值越小,表明误差显著水平越低,模型拟合越显著。
1.2.3 种群生长动态三维模拟 整理野外调研数据,结合种群生长因子关系模型,分别对各样地黄牛木个体的树高、冠幅以及数量进行测算,利用ArcGIS 的Arcsence 三维图形处理功能,分别绘制各样地现阶段、5 径阶、10 径阶、20 径阶后黄牛木种群的三维模拟图,直观的呈现种群生长变化的动态规律。
2 结果与分析
2.1 种群生长因子统计分析
种群生长因子信息是研究种群生态学的重要基础数据,整理外业调查数据,统计黄牛木种群生长因子信息,结果见表1。
从表1 的数据可以看出,茶山样地黄牛木植株为788 株,最高径阶个体的胸径为19.5 cm;白云山样地黄牛木植株为956 株,最高径阶个体的胸径为16.6 cm;石花山样地黄牛木植株为160株,最高径阶个体的胸径为16.1 cm。其中,茶山和白云山样地的黄牛木幼树和小树占种群总数的70%以上(茶山606 株,占比为76.9 %;白云山742 株,占比为77.6%),明显高于中树和大树的植树数量(茶山175 株,占比为22.2 %;白云山210 株,占比为22.0%),老树的植株数量均不及种群总数的1%(茶山7 株,白云山4 株),种群生长呈现“金字塔型”,表明这两个样地的黄牛木种群处于生长旺盛期,为增长型种群。而在石花山样地中,黄牛木幼树与小树共计70 株(占比为43.7%),中树和大树共计89 株(占比为55.6%),老树仅有1 株(占比为0.63%),种群生长呈现“纺锤型”,表明该样地的黄牛木种群生长趋于稳定,为稳定型种群。
从表1 的数据还可以看出,茶山样地中各径阶数量随径阶的增加而减少,胸径、树高、冠幅等林木生长因子均呈现正常生长状态,说明该黄牛木群落未受到过多的环境或人为破坏,生境比较稳定。白云山样地表现为种群数量随径阶增大先增加后减少,在第2 径阶(1 ≤D<3 cm)达到最大值352 株,其他林木生长因子均呈现正常生长状态,说明该样地生境相对稳定。与另外两个样地不同,石花山样地的调查面积稍小,但种群数量却明显偏少,不及其他两个样地的一半,种群数量变化趋势与白云山有相似之处,但峰值出现时间不同,即种群数量随径阶增大先增加后减少,在第4 径阶达到峰值,查阅调查记录发现,石花山样地的植被丰富度明显高于茶山样地和白云山样地,不难想象该样地中黄牛木种群的生长压力大于其他两个样地。此外,石花山样地第8径阶(13 ≤D<15 cm)的苗木高度(2.3 m)和冠幅(0.1 m)存在异常,两者的值均比相邻径阶的小,这与植物的正常生长规律相悖。查阅调查记录发现,该样地中的所有植株,第8 径阶生长异常的仅有此一株,且四周均为其他高大的乔木,阻碍了该株黄牛木的增高和冠幅伸展,说明该样地的黄牛木种群在生长发育过程中受到了一定的干扰。
定植时将秧苗带育苗营养块定植到本田定植穴内,一定要及时浇灌定植水,促进根系快速萌发伸长,防止因缺水使外露根系受到损伤。待水分渗漏以后,用土将幼苗基部及营养块表面培土覆盖2~4厘米。
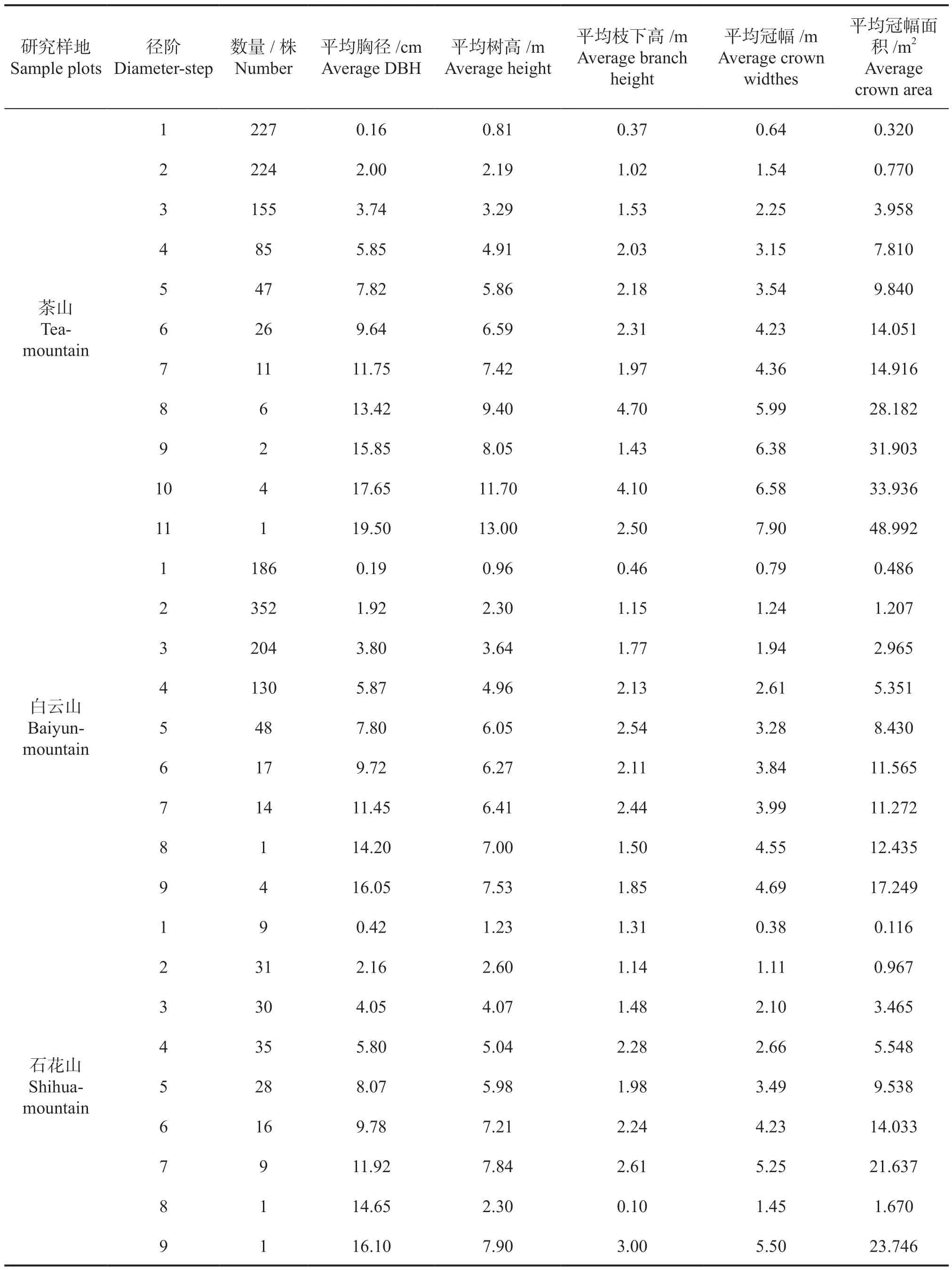
表1 黄牛木种群生长因子统计结果Tab.1 The statistical results with growth factor of Cratoxylum cochinchinense population
2.2 种群生长因子关系模型构建
胸径、树高、冠幅是林分调查中最重要的生长因子,一般来说,林木个体胸径越大,树高和冠幅的值越大,即林分的树高、冠幅与胸径之间存在正相关关系。为进一步探讨黄牛木个体生长因子之间的关系特征,掌握树高、冠幅随胸径变化的动态规律,将3 个样地的种群数量作平滑处理后加权平均,其余生长因子合并后加权平均,结果见表2。
从表2 可以看出,黄牛木种群的个体数量随径阶增大先增加后减少,在第2 径阶达到峰值,根据种群生长初期的数量分布特征,认为这属于正常现象。种群的平均树高、平均枝下高、平均冠幅、平均冠幅面积等均随径阶的增加而增大,符合林木生长的一般规律。
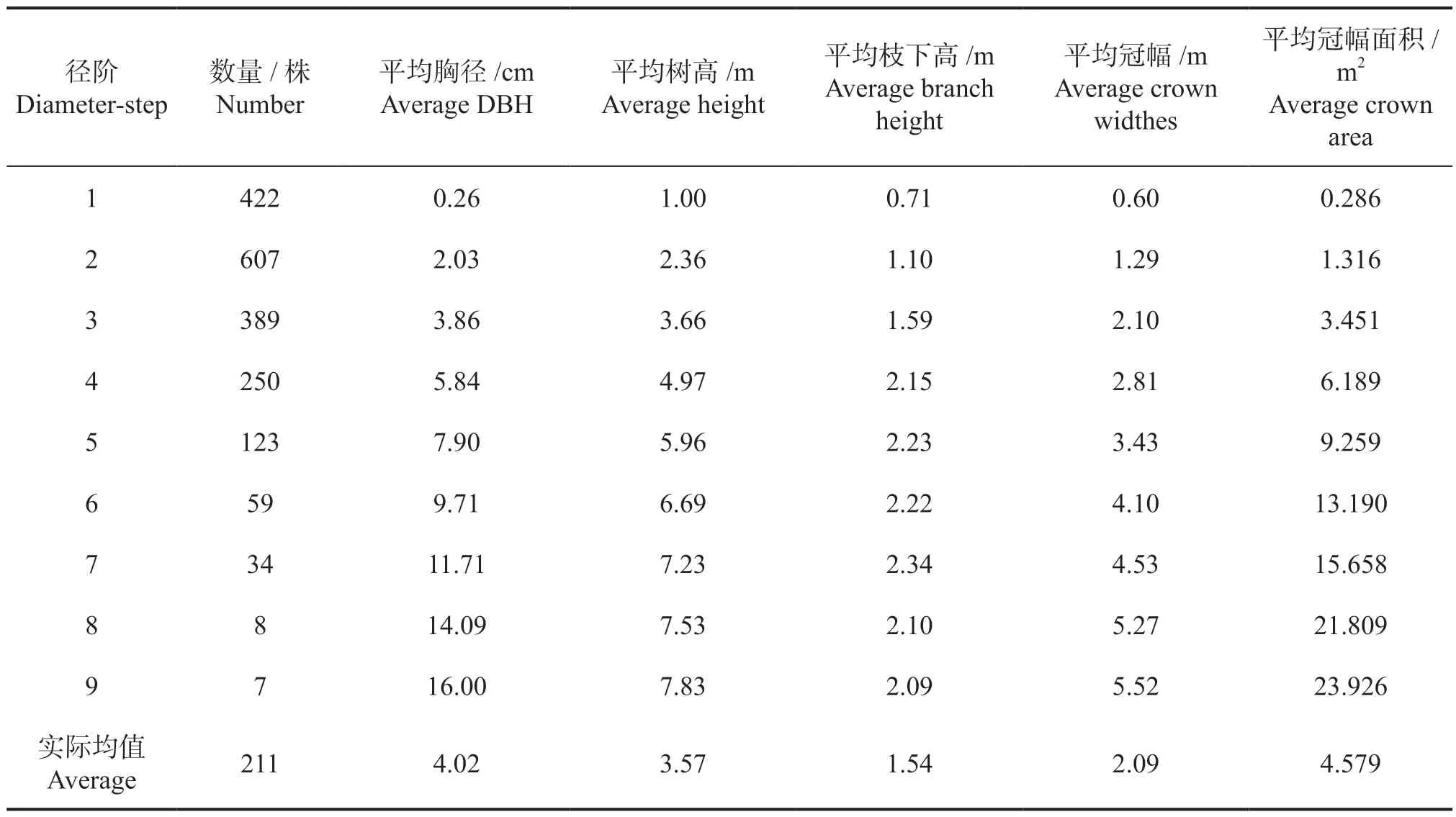
表2 黄牛木种群个体生长因子处理结果Tab.2 The processing results with growth factor of Cratoxylum cochinchinense population
将表2 的数据导入SPSS 对林木生长因子进行回归分析,通过比较拟合模型的各项参数值,结合林木的生物学特性选取最佳拟合模型。黄牛木的树高与胸径、冠幅与胸径的拟合曲线相关模型参数估计值(表3~4)。
从表3 可知,树高和胸径显著正相关,各个方程拟合情况良好,其中,二次函数的拟合效果最佳,其R2值0.998 明显高于其他函数,P值小于0.000,回归模型系数为0.999,F值为1 551.468;幂函数次之。但从生物学特性方面考虑,二次函数模型适用于林分的蓄积量、生物量等,不适用于林木个体拟合,相比之下,幂函数拟合模型更符合林木生长规律,其函数公式为y1=1.903x0.525(R2=0.988,P<0.000),回归模型系数为0.994,F值为598.685。通过调查数据验证模拟函数的精确度,将黄牛木种群的实际平均胸径4.02 cm 代入回归方程,计算得到模拟树高为3.95 m,与实际调查数据3.57 m 稍有差距,尚属正常误差范围。
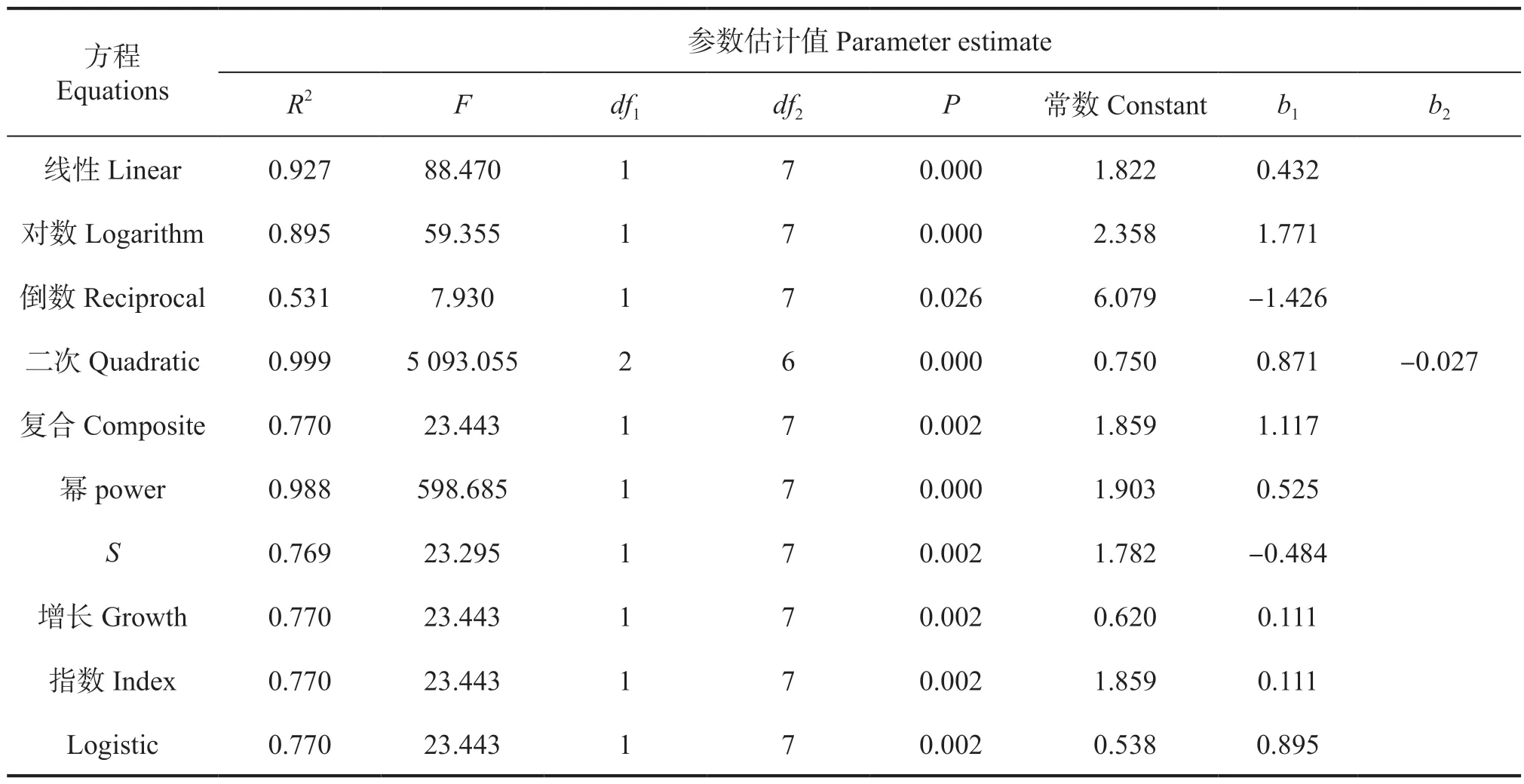
表3 黄牛木种群树高-胸径模型汇总和参数估计值Tab.3 The functional model equation and parameter estimation with Height - DBH of Cratoxylum cochinchinense population
从表4 可知,冠幅和胸径显著正相关,各个方程拟合情况良好。同样地,考虑满足林木生物学特性和实际生长规律,最终认为幂函数为冠幅与胸径的最佳拟合模型,函数公式为y2=1.1x0.558(R2=0.975,P<0.000),回归模型系数为0.987,F值为270.836。根据现场调查数据,黄牛木的实际平均胸径为4.02 cm,由回归方程计算出黄牛木的模拟冠幅为2.39 m,近似于实际调查数据2.09 m。
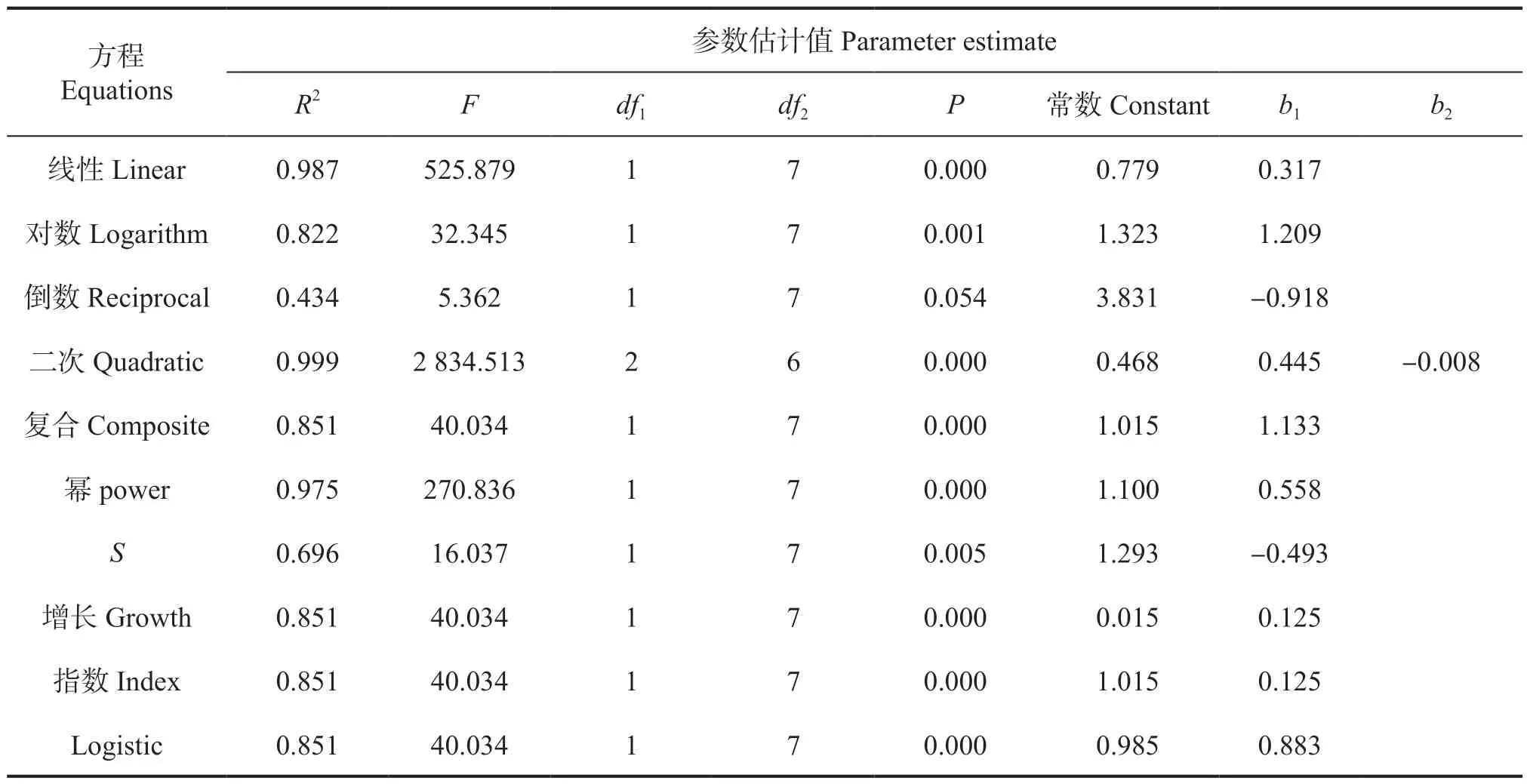
表4 黄牛木种群冠幅-胸径模型汇总和参数估计值Tab.4 The functional model equation and parameter estimation with Crown width - DBH of Cratoxylum cochinchinense population
2.3 种群数量变化动态分析
数量变化动态是种群数量特征研究的重要内容,将3 个样地的种群数量作平滑处理后加权平均,具体数值详见表2。参照上文树高-胸径、冠幅-胸径函数模型的构建方式,将黄牛木种群数量与胸径进行拟合分析,通过比较数量与胸径关系拟合曲线相关模型参数估计值(表5),得到数量-胸径的最佳拟合模型为指数函数,函数公式为:y3=2 088.9e-0.641x(R2=0 .965,P<0.000),回归模型系数为0.982,F值为227.030。
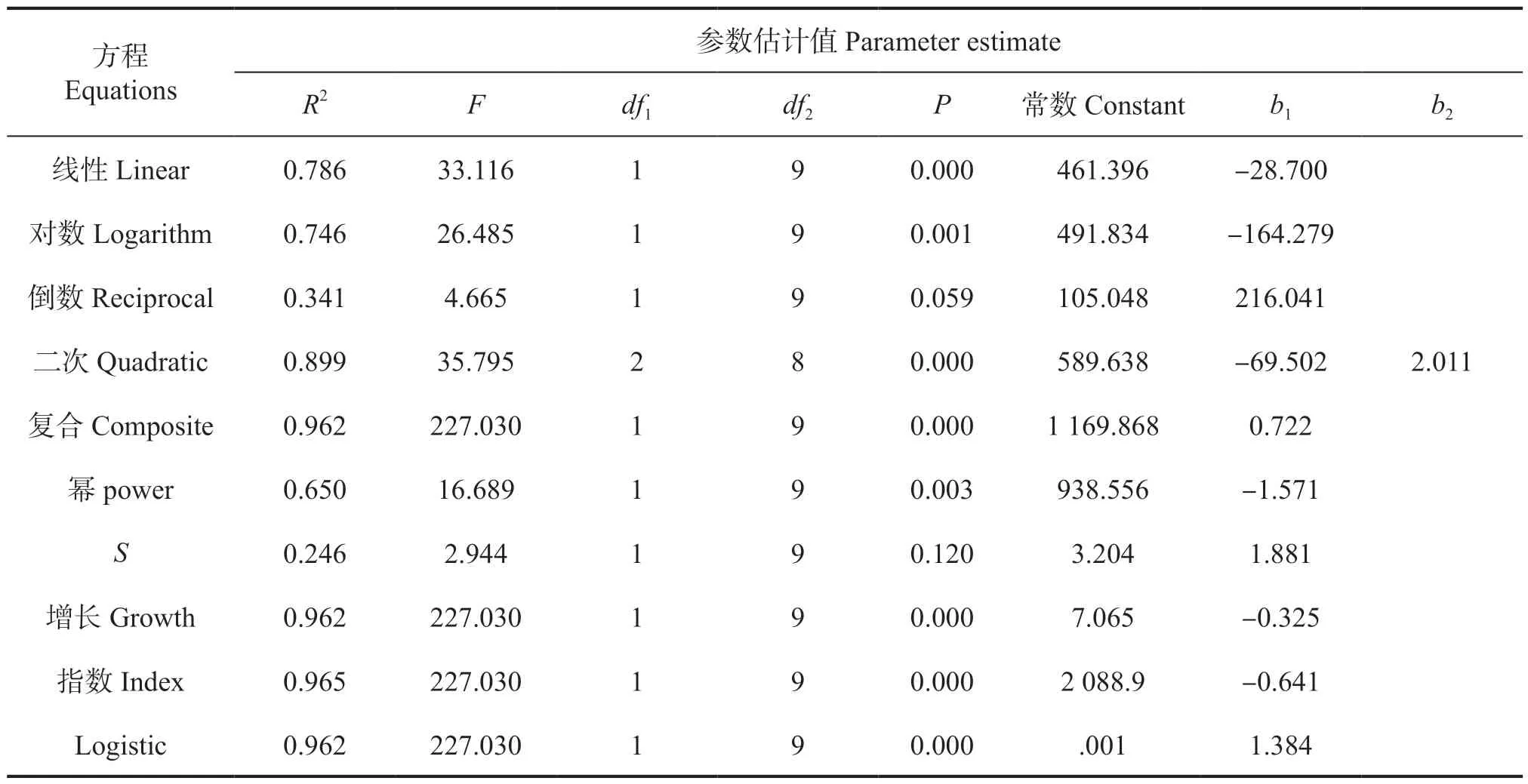
表5 黄牛木种群数量-胸径模型汇总和参数估计值Tab.5 The functional model equation and parameter estimation with Number - DBH of Cratoxylum cochinchinense population
为进一步探究黄牛木种群数量的变化规律,参照时间序列预测法在相关研究中的运用[21],利用一次平均推移法预测第Ⅱ径级、第Ⅲ径级、第Ⅳ径级、第Ⅴ径级、第Ⅵ径级、第Ⅶ径级后的种群数量,结果如图1。从图1 可以看出,黄牛木种群各径阶数量峰值在预测序列中依次向后推移,个体数量随时间推移逐渐向中老径阶集中,老龄个体增多,幼龄个体减少,幼龄株数更新不足,最终老龄株数也将呈现减少的衰退势态。
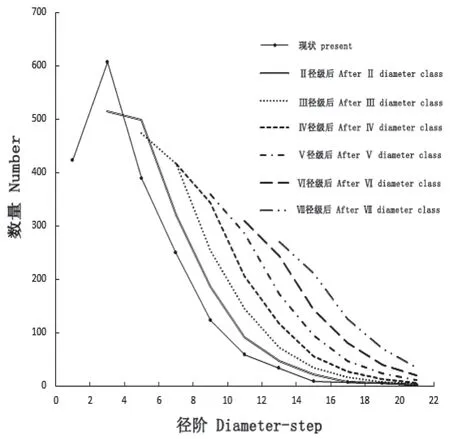
图1 黄牛木种群各径阶数量变化预测Fig.1 The quantity prediction of different diameterstep of Cratoxylum cochinchinense population
2.4 种群生长动态三维模拟
种群生长三维图的构建,几乎是对其空间分布和生长动态情况的真实再现。将调查及模拟数据导入ArcScene,分别生成各样地黄牛木种群不同生长阶段的三维模拟图,结果见图2-5。
从图2 可以看出,现阶段黄牛木大树较少,幼树和小树数量占比较大,种群覆盖率较低,冠层呈现少量重叠覆盖现象。
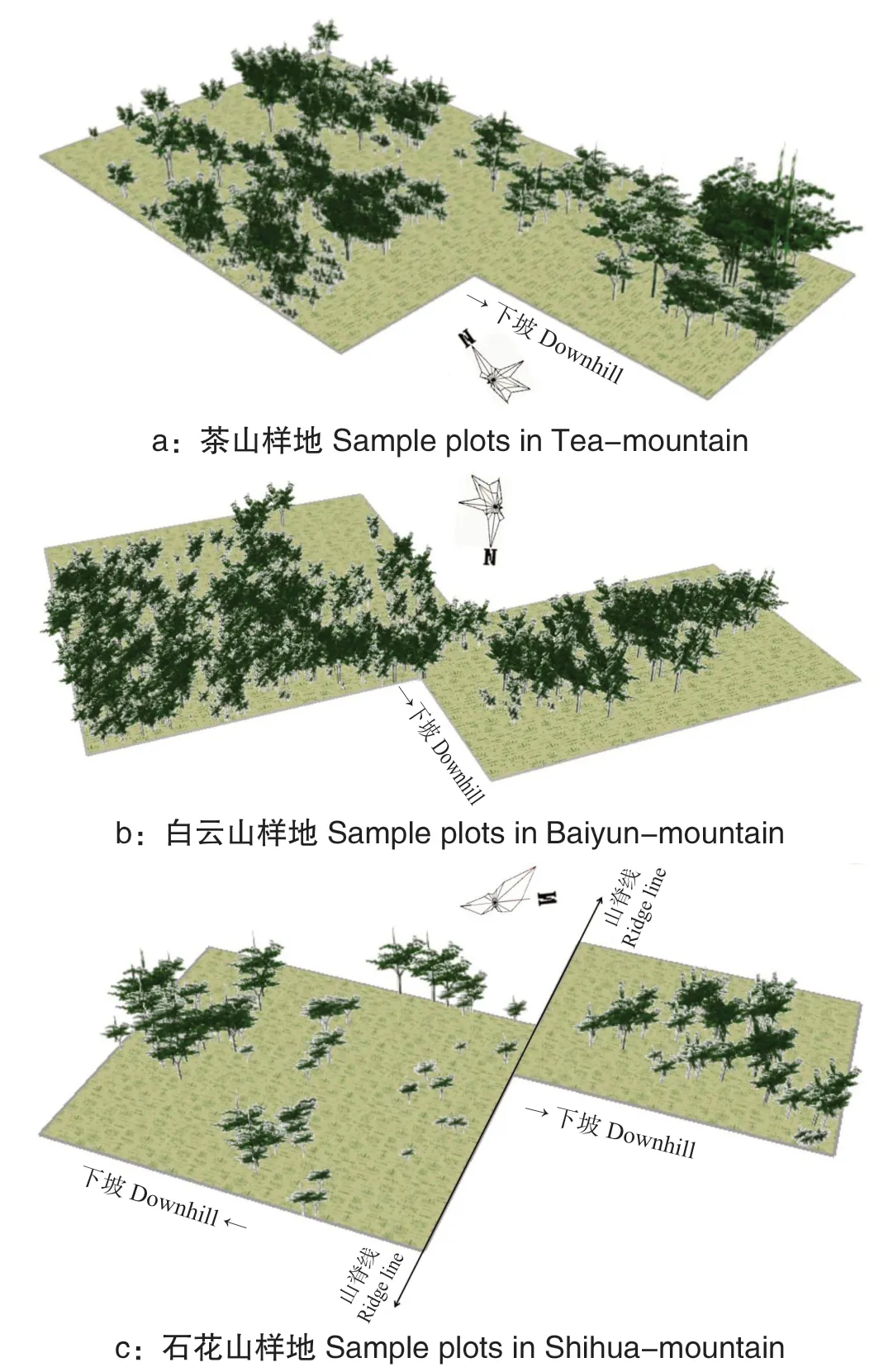
图2 黄牛木种群生长现状Fig.2 The map with growth status of Cratoxylum cochinchinense population
从图3 可以看出,经过5 径阶的生长,黄牛木种群冠层相互重叠覆盖程度愈发明显,冠层覆盖率提高,种群内部斗争将逐渐加剧,种群生长速度也将逐步下降,此时应考虑对种群进行适当疏伐。
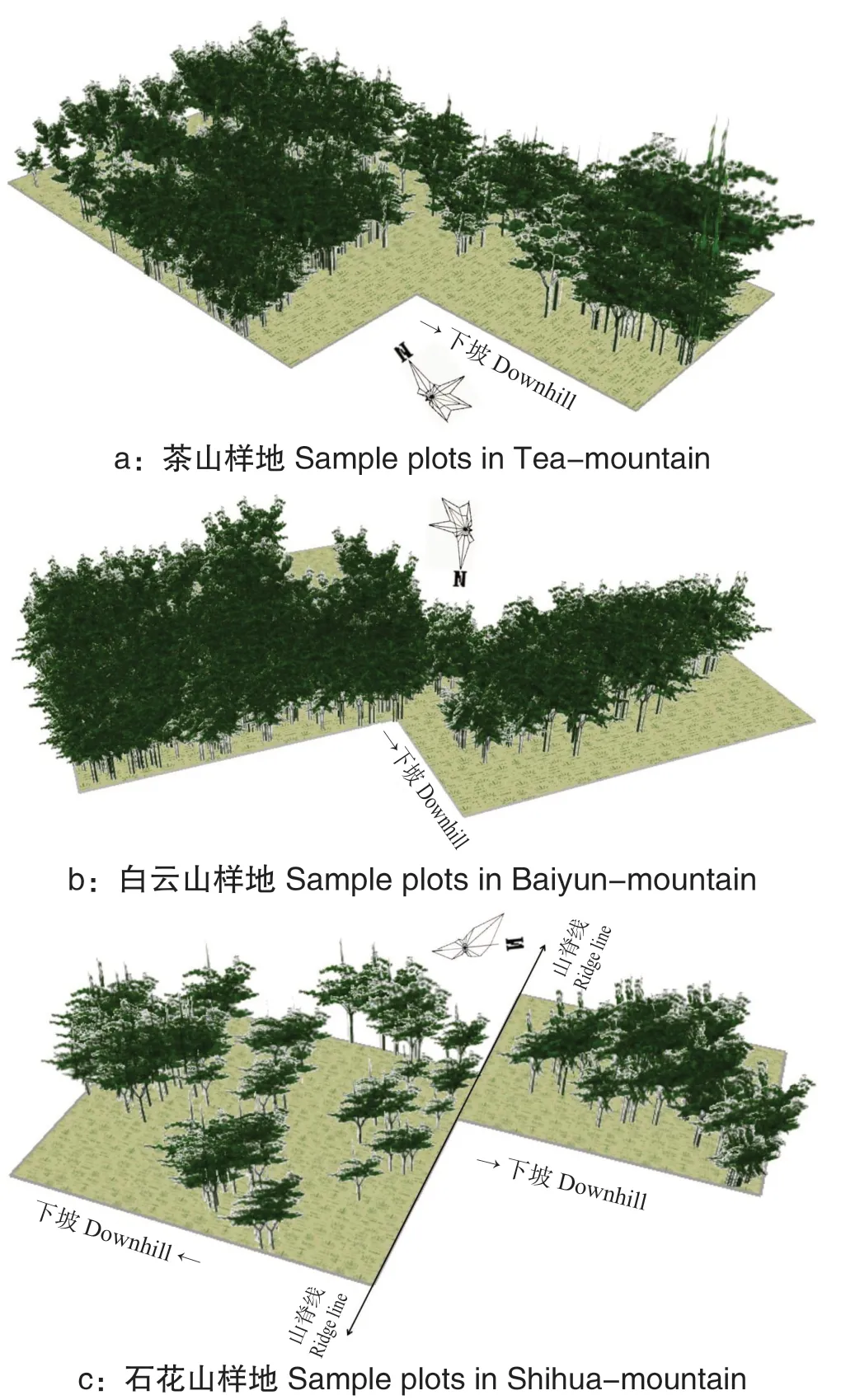
图3 黄牛木种群5 径阶后生长模拟Fig.3 The map with growth simulation after 5 diameter-step of Cratoxylum cochinchinense population
从图4 可以看出,种群经过上一轮的种内斗争,形成强者更强、弱者更弱的局面,种群的扩散及生长总体呈现上升趋势,但不明显。此时应适当伐除样地内老、弱、病、残植株以及部分幼苗,使得正常生长的黄牛木个体拥有充足的养分,确保种群持续增长。
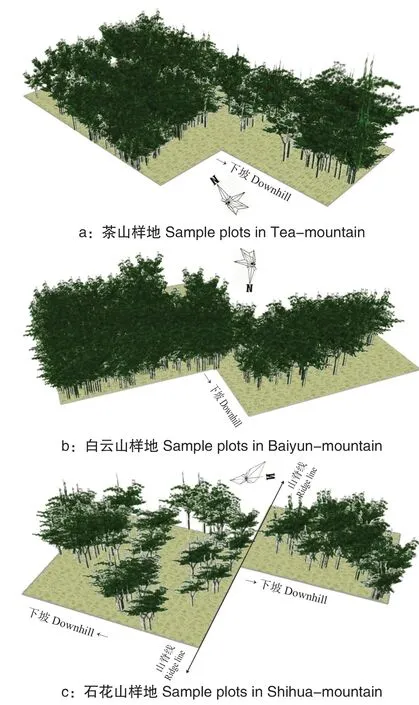
图4 黄牛木种群10 径阶后生长模拟Fig.4 The map with growth simulation after 10 diameter-step of Cratoxylum cochinchinense population
从图5 可看出,20 径阶后,各样地黄牛木的生长已有显著不同,几乎全部植株已长成大树,小树明显减少,种群冠层覆盖程度明显上升,绝大部分树木进入生理衰老期。此时应该加强抚育管理,加大间伐力度,从而促进黄牛木种群的演替与更新。
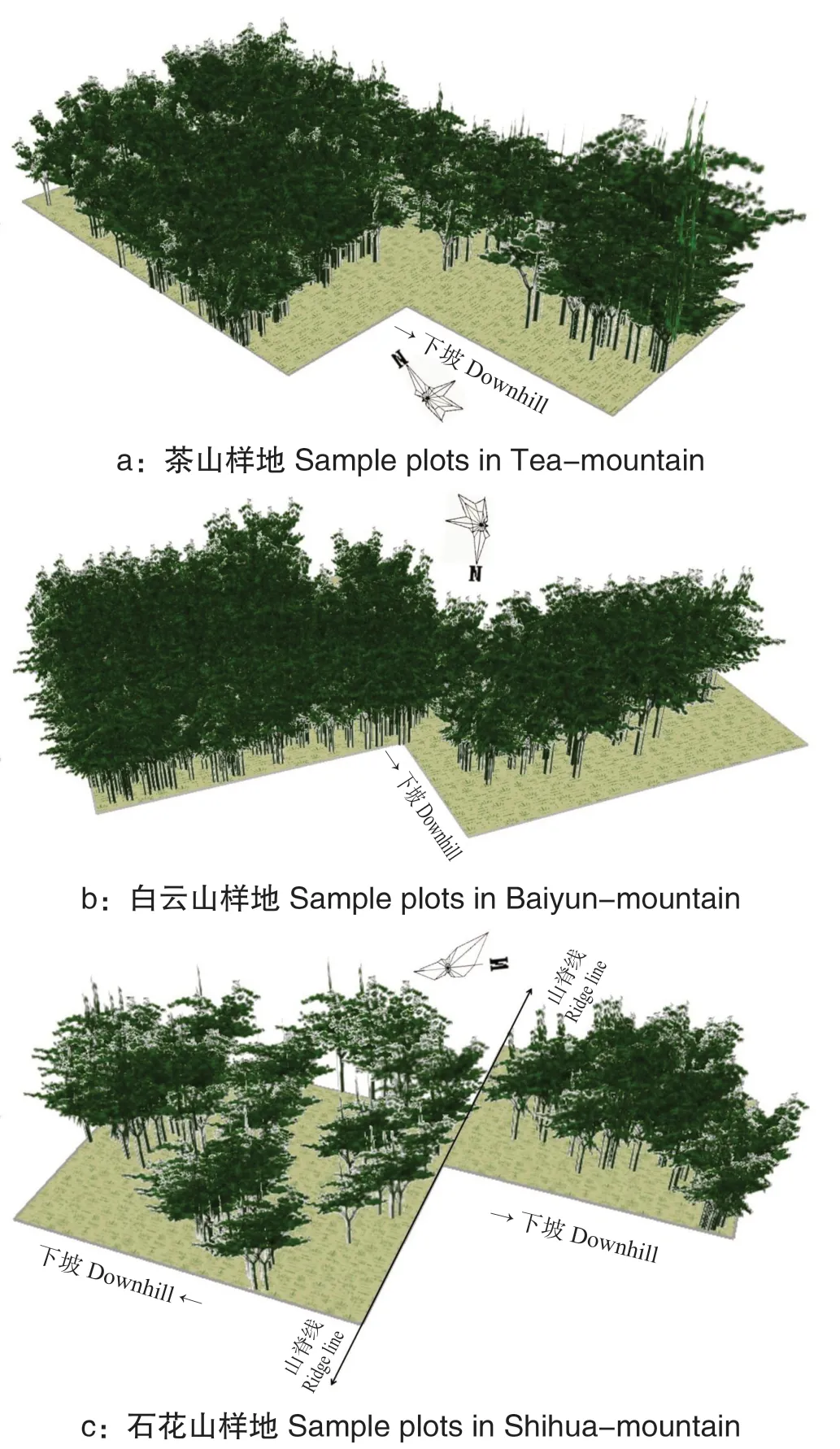
图5 黄牛木种群20 径阶后生长模拟Fig.5 The map with growth simulation after 20 diameterstep of Cratoxylum cochinchinense population
3 结论与讨论
调研数据显示,除石花山的个别生长阶段外,黄牛木种群的胸径、树高、冠幅等生长因子基本处于正常生长状态。在种群生长过程中,白云山和石花山种群数量先增加后减少(分别在第2 径阶和第4 径阶达到峰值),原因可能是幼树在萌发或生长过程中受到干扰,如种子成熟时被动物过分取食、雨水过多或过少导致种子发芽率降低、幼树生长期受到人为破坏等均会造成幼树数量的减少;石花山丰富的群落植被尤其是茂密的林下灌木和密实的芒萁草本层也可能是黄牛木幼苗萌发生长的主要限制因素,具体原因还有待进一步查证。
树高、冠幅、胸径是衡量林分生长重要的指标[22],对于生长因子之间相互关系的函数模型,学术界已有不少研究[23-25]。胸径测定简单方便、精度高,而树高、冠幅的测定却费时耗工,误差无法控制,这给林业生产与实践带来较大困难。本研究构建黄牛木树高-胸径、冠幅-胸径函数模型有较好的生物学意义,一方面为野外调查时树高、冠幅的测定提供了新的思路,另一方面为该地区黄牛木天然林的生长预测以及抚育管理提供理论依据。但值得注意的是,模型的构建是通过有限的空间和数据分析而得,具有一定的理想性和局限性,在实际工作中,还要结合林木的生长环境进行综合分析,从而确定种群的最佳间伐时间和抚育措施。
种群数量与胸径的函数模型反映了种群生长变化规律,可为预测种群动态及适时疏伐提供理论依据,此前也有类似研究[26-27]。种群的演替更新是种群生物学特性与环境因素共同作用的结果[28-29]。本研究运用一次平均推移法预测黄牛木种群的更新动态,结果为黄牛木种群数量随时间推移逐渐向中老径阶集中,老龄个体增多,幼龄个体减少,与前人的研究结论一致[30-31]。可以推断,由于缺乏可更新的幼龄个体,如不采取适当的护林抚育措施,当地黄牛木种群将趋于衰退,种群稳定性长期维持较为困难。
树种的生长处于不断变化中,种群生长三维模拟图反映的是理想状态下黄牛木的生长规律,但也在很大程度上反映了林分的生长现状和动态规律,有利于实现黄牛木天然林的最大景观价值和森林生产效益。从人工经营管理的角度而言,掌握黄牛木的生长规律和变化动态,有利于引导管理者适时开展合理的疏伐工作,实现种苗培育和营林保护的效益最大化,不仅为评价林木生境与森林生长优劣程度提供参考,同时为各级林业工作部门制定相关政策方针提供理论基础,促进我国林业产业的可持续发展[32]。
猜你喜欢
安徽农业科学(2022年19期)2022-10-29
绿色天府(2022年2期)2022-03-16
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
林业科技情报(2021年3期)2021-09-01
林业科技情报(2021年2期)2021-07-13
新教育(2018年27期)2019-01-08
中南林业调查规划(2017年1期)2017-12-19
中国卫生(2016年3期)2016-11-12
创业家(2015年2期)2015-02-27
