
大青山米老排树干径向生长变化及其对环境因子的响应
2020-02-12 09:02:46杨保国刘士玲庞圣江韦菊玲王庆灵
中南林业科技大学学报 2020年2期
杨保国,刘士玲,庞圣江,张 培,郝 建,韦菊玲,王庆灵,
邹位锡1,2,覃 瑶1,2
(1.中国林业科学研究院热带林业实验中心,广西 凭祥 532699; 2.广西友谊关森林生态系统国家定位观测研究站,广西 凭祥 532699)
影响树木生长主要有两方面因素:生物学遗传特性和生态因子,此外还受林木竞争、人工抚育措施等诸多因子影响[1-2]。树干径向生长是研究树木生长的重要途径,尤其是树木径向生长与气象因子的关系研究,对揭示树木生长对气候条件的适应性很有意义[3]。目前利用树木年轮-气候资料研究树木生长机制的方法已较为成熟[4-5],然而,作为生命体的树木,其径向生长与气候因子间存在着十分复杂的作用关系,而且这种作用关系受到区域气候条件和树木本身生理特性等多方面因素的综合影响,在不同个体及空间和时间上也会表现出一定的差异。但传统年轮气候研究对径向生长-气候响应的生理机制时间分辨率较粗,不能体现出树木径向生长在小时或者更短的时间尺度上的变化规律。所以自20 世纪70年代以来,国外开始广泛使用树木径向变化测量仪(Dendrometer)来研究树干的径向生长过程[6-9],国内亦有许多学者进行此类研究,但主要集中在我国北方地区[10-13], 利用径向变化测量仪对南方乡土阔叶树种径向生长变化在日尺度、甚至更小尺度上的研究较少[14]。这些研究证实,从小时或更短的时间尺度上研究树干径向生长,对从生理生态学角度深入认识树木生长—环境因子响应机理具有重要价值[15]。
米老排Mytilaria laosensis是南亚热带区域适生范围广的速生乡土珍贵用材树种,郭文福[16]等研究表明5月和9月是米老排生长季的两个高峰期。本研究基于树木径向测量和气象因子同步观测,对南亚热带大青山林区米老排第一个生长高峰期5月的径向微生长进行监测研究,试图揭示不同分化等级米老排径向日变化规律、不同天气条件下生长变化及径向生长-环境因子的关系,从而为理解米老排生长-气候响应的生理机制、森林培育和保护等提供一定的基础数据。
1 研究区概况
研究区域位于广西壮族自治区友谊关森林生态系统国家定位观测研究站所在的大青山林区(106°41′~106°59′E,21°57′~22°16′N),属南亚热带湿润—半湿润季风气候;年降水量1 200 ~ 1 500 mm,4—10月份为雨季,占全年降水量80%;年蒸发量1 261 ~1 388 mm,年均气温 21.6 ℃,≥10 ℃活动积温6 000 ~7 600 ℃,相对湿度80% ~84%,海拔430 ~680 m。土壤为由花岗岩发育而成的赤红壤,土层厚度100 ~ 150 cm,腐殖质层厚度5 ~10 cm。
2 材料与方法
2018年1月份选择生长良好的米老排人工林,建立1 个20 m×20 m 的标准地,其海拔高度为500 m,坡向东北,坡位中部。根据林木生长情况,在林分内选取优势木、平均木、被压木[17]共3株样木,在树干1.3 m 高处刮去部分翘裂及干枯树皮,安装点状径向生长测量仪(Dendrometer,DC3 型,Ecomati 公 司, 分 辨 率1 μm, 误 差±3.3 μm,连续记录径向生长变化数据)。样木基本信息如下(表1)。

表1 研究样木基本信息Table 1 Basic information of the sample trees
同步的气象数据采用位于热带林业实验中心伏波实验场的友谊关生态定位站红星气象站数据(直线距离<1.5 km),气象环境要素包括气温(℃)、降水(mm)、辐射(μmol·s-1·m-2)、空气湿度(%)和主要根系层20 cm 土壤体积含水率(%)等因子。所有仪器步幅设定均为15 min。
本研究仅以5月份检测记录的径向数据和气象数据进行日尺度的初步分析。数据用Excel 进行初步整理分析,用SPSS19.0、SigmaPlot10.0 进行数据相关分析、绘图。
3 结果与分析
3.1 米老排径生长日变化特征
为了精确判读树木直径生长及其时间分布,国际上通常人为地将日变化划分为3 个阶段:收缩阶段、膨胀阶段、生长阶段[6,10-11,18]。结果表明:5月份米老排径向生长日变化趋势呈“U”型(图1),不同分化等级间树干径向生长量不同,且差异达极显著水平(P<0.01)。
根据以上阶段划分,计算了观测期内树干径向日变化各个阶段的起始时间范围、各阶段平均持续时间以及各阶段径向生长变化量(表2)。一般来说,从08:00—09:30 开始,由于树木冠层的蒸腾作用增强,树木直径进入收缩阶段,不同分化等级的平均起始时间表现为平均木(08:50)略早于优势木(09:00),被压木(09:35)最晚。从16:30—15:30开始,由于蒸腾作用逐渐减弱,树木从根部吸水补充,树干不断膨胀,其直径进入膨胀阶段,平均木(16:30)略早于被压木(17:00),优势木(17:10)最晚。生长阶段一般发生在夜间至翌日凌晨。

图1 米老排5月份累计径向生长平均日变化Fig.1 Cumulative daily radial growth of Mytilaria laosensis in May

表2 不同分化等级米老排树干径向日变化†Table 2 Daily change of stem radial growth of Mytilaria laosensis with different differentiation classes
3.2 米老排径向生长变化与环境因子相关分析
米老排径向生长与环境因子监测资料分析结果表明(图2 ~5),不同分化等级米老排径向生长与环境因子的相关趋势基本一致,从Pearson 相关系数来看,总体表现为与相对空气湿度、降水量呈极显著正相关关系,与气温、总辐射呈极显著负相关关系,而20 cm 土壤含水率则表现为与优势木径向生长负相关,与平均木、被压木正相关,且均无显著性相关。进一步对米老排径向生长瞬时监测数据与环境因子(见表3,提前1 h、即时、滞后1 h)相关性分析表明,各因子普遍存在一定的滞后效应,总辐射、降水量滞后0 ~0.5 h, 气温、相对空气湿度则存在0.5 ~3 h 滞后效应,20 cm 土壤含水率的滞后效应甚至可达8 h 以上。不同分化等级米老排与环境因子的相关性也有所不同,总体来说,平均木>优势木>被压木。
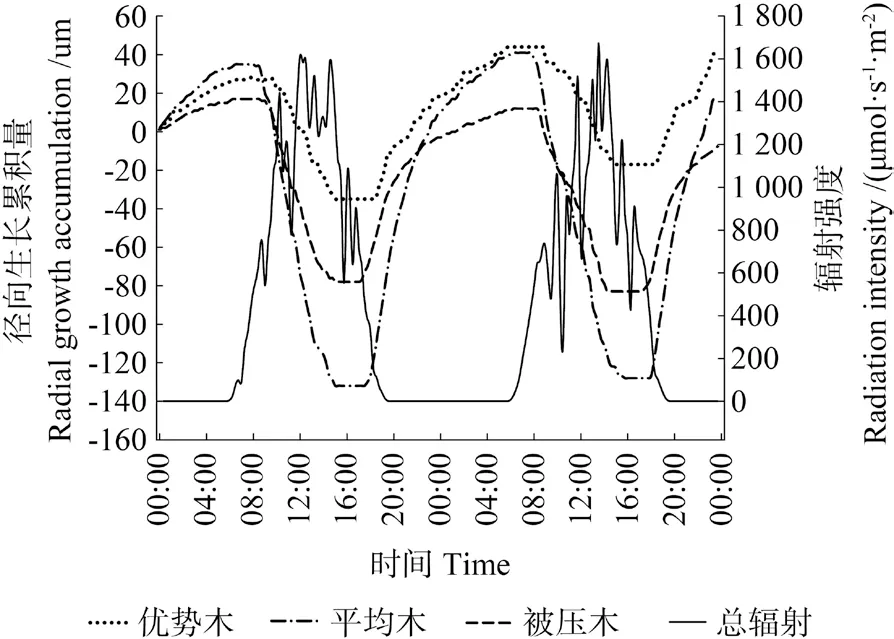
图2 晴天条件下米老排树干直径变化Fig.2 Daily change of stem diameter of Mytilaria laosensis in sunny day
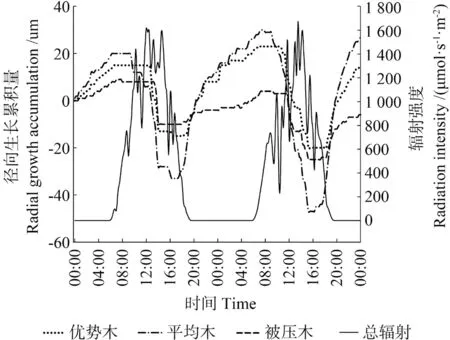
图3 阴天条件下米老排树干直径变化Fig.3 Daily change of stem diameter of Mytilaria laosensis in cloudy day
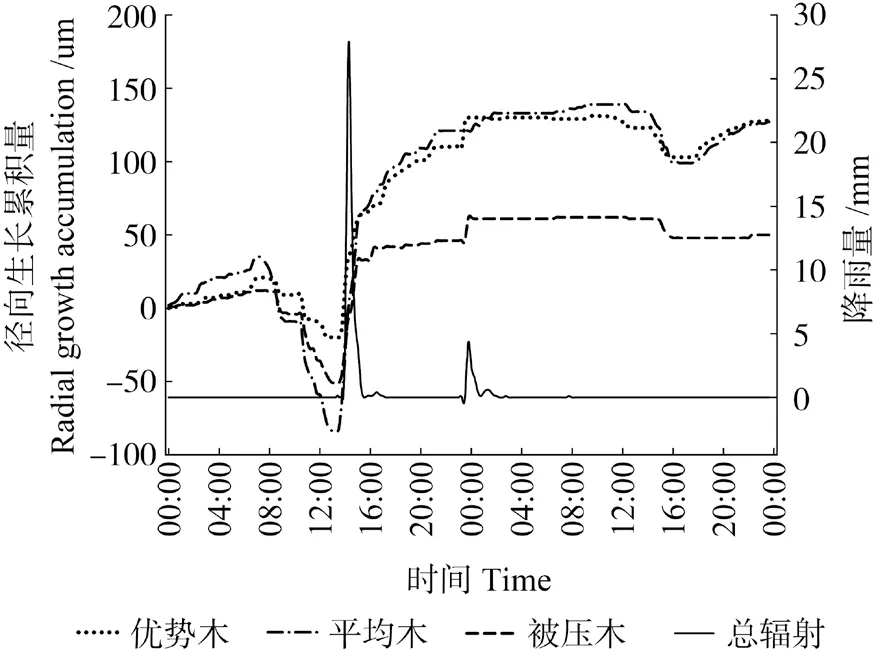
图4 雨天条件下米老排树干直径变化Fig.4 Daily change of stem diameter of Mytilaria laosensis in raniy day
4 结论与讨论
4.1 米老排个体径向生长日变化差异分析

表3 米老排径向生长变化与环境因子即时及滞后效应相关统计†Table 3 Correlation statistics of immediate and lag effects of radial growth change and environmental factors
树干径向变化可分为不可逆和可逆两个过程,前者是指树干形成层细胞的分裂和伸长而产生韧皮部、木质部形成过程[19];后者是主要指树体水分收支平衡所引起的树干体积物理涨缩变化[20-21], 此外,温度等微气象的变化也可能通过影响树干体积的热胀冷缩或细胞生长影响树干径向变化[13,22]。不同分化等级米老排径向生长日变化的3 个阶段的起始时间、持续时间、变化量有较大差异,有些日变化由于在膨胀阶段不能恢复到前一日的最大值,从而出现零生长或负生长,甚至有些林木个体在监测期间径向变化呈现负增长趋势,这主要由微气象、环境因子及树木本身个体差异造成的,树木个体差异在径向变化中的差异主要体现在生长阶段(表4),其中冠幅厚度与生长阶段变化量的Pearson 相关性达到显著水平(P<0.05),这主要是因为冠幅厚度越大,叶片数越多,可以占有更多的光照、吸收更多的水分,合成更多的光合产物,进而利于径向生长,管伟等[23]的研究也认为能够获取更多水热资源的优势度大的树木生长更快。有些学者对树木径向生长日变化的研究中虽然也出现零生长或负生长现象[11,23],但尚未发现在生长季监测期间林木径向呈缩小现象,本研究中米老排被压木径向呈缩小趋势,可能因为该林分已完全郁闭,米老排又属强阳性树种,被压木在林分下层,竞争力较弱,长期的不到充足的光、水、肥,生活力减弱,生长受到压制,日趋枯死[24]。

表4 不同阶段径向日变化与林木特征Pearson 相关分析†Table 4 Pearson correlation analysis of radial daily variation and tree characteristics in different stages
4.2 米老排径生长变化对天气条件响应
雨天时米老排径向日净生长量远大于晴天、阴天条件下,呈持续增长趋势,这种净生长量主要来源两个方面,一是雨天时,树干及树皮吸水充分膨胀,且土壤可以充分供水,蒸腾减弱,树干失水量明显减少,所以树干的失水-补水过程微弱[25];另一方面是树木自身的生长量,但前者的变化量远超后者,完全掩盖了树木自身的生长量,这也是导致雨天时3 种分化等级米老排径向日净生长量差异不显著的主要原因,这与管伟[25]对华北落叶松Larix principis-rupprechtii的研究结果一致。但是,降水后的几天内,由于树干、树皮水分变化剧烈,树木径向生长可能会出现“负生长”现象。晴天条件下光照充足,光合作用产生的光合产物较阴天时更多,因此晴天条件米老排下径向日净生长量要略高于阴天条件下。从日生长量变化幅度来看,雨天变化量最大,这主要受降水影响,树皮、树干吸水膨胀所导致,晴天> 阴天,则主要受林木体内失水-补水过程影响,显然晴天条件下这个过程表现更为强烈。虽然平均木日生长量变幅总是最大,但累计生长量却是优势木最大,这说明平均木径向变化受植物体内水分状况影响较大。被压木径向生长趋于减小,可能是因为其体内水分“入不敷出”,即根部受周围树木根系挤压,其吸水能力日益减弱,植物体吸收的水分不足以提供其消耗(蒸腾、生长),所以在阴天和雨天时其径生长变幅较小。
4.3 米老排径向生长变化的影响因子
树木径向生长与降水量、相对空气湿度正相关,与气温、总辐射负相关,且均存在不同程度的滞后性,这与大多数学者的研究结果一 致[3,8,10,14,23,26];但土壤含水率与径向生长未表现出显著的相关性,这与肖生春[11]的研究结果有所不同,可能是树种特性、区域气候、立地条件等差异太大所导致。降水一方面会导致林木吸涨,一方面通过增加土壤水分来控制林木径向生长[21]。相对空气湿度、气温、总辐射主要通过影响林木蒸腾量,进而影响林木树干液流,最终控制径向生长变化[6,27-28];相对空气湿度高,气温和总辐射低,林木蒸腾弱,树干液流小,根部持续吸水,林木吸水膨胀;反之,则林木蒸腾强,树干液流大,根部吸水主要用于蒸腾,林木径向变化小。此外,温度也通过影响林木本身光合能力和改变土壤含水率、水势等环境因子,影响林木的径向生长[13,29]。Zweifel 等[30]研究认为土壤含水率主要通过影响林木茎干组织水分,改变细胞伸长生长的主要动力——形成层细胞膨压,最终影响到林木的径向生长。不同分化等级样木由于胸径、树高、冠幅、叶片数、根部吸水能力有所差异,导致其径向生长变化与环境因子相关性有所不同。
综上所述,不同分化等级米老排由于林木自身胸径、树高、冠幅厚度、叶片数、根系吸水能力等的差异,导致其受到环境因子影响程度的不同,最终影响到其在径向生长日变化量、不同天气条件下的变化量以及对环境因子的响应程度有所差异。本研究筛选了气温、相对空气湿度、总辐射、降水量、土壤含水率5 个影响因子与不同分化等级米老排径向生长进行统计相关分析,以便直观了解不同分化等级林木径向生长与环境因子的相互关系。但受限于仅选取该区域5月份的3株米老排径向生长和环境因子数据进行分析,该结论需要进一步验证。对于米老排径向生长月变化、年变化以及更深层次的影响机制,还需要长时间观测以及引入更多的影响因子(林木特性、林分结构、环境因子等)做进一步分析。
猜你喜欢
绿色科技(2021年21期)2021-11-26 09:12:06
军事文摘(2021年16期)2021-11-05 08:49:16
内蒙古林业调查设计(2021年3期)2021-07-06 14:12:00
现代园艺(2018年1期)2018-03-15 07:56:13
现代园艺(2017年23期)2018-01-18 06:58:17
现代园艺(2017年21期)2018-01-03 06:42:15
小猕猴学习画刊(2017年1期)2017-02-17 16:04:14
小猕猴学习画刊(2017年1期)2017-02-17 15:57:35
广东第二课堂·小学(2016年11期)2016-12-06 14:29:33
新农业(2016年17期)2016-08-16 12:02:38
