
松材线虫扩散型四龄幼虫向成虫转型发育阶段研究
2020-01-18 07:21张伟理永霞刘振凯高梦鸽孟繁丽张星耀
中国森林病虫 2020年1期
张伟理永霞刘振凯高梦鸽孟繁丽张星耀
(1.中国林业科学研究院林业新技术研究所,北京 100091;2.南京林业大学南方现代林业协同创新中心,江苏南京 210037)
松材线虫病(Pine wilt disease)即松树萎蔫病,是以松材线虫Bursaphelenchus xylophilus为病原[1],墨天牛属昆虫Monochamusspp.为主要媒介[2],综合人为参与、寄主松树、相关伴生菌和环境因素互作的一种复杂病害系统[3-4]。该病于20 世纪80 年代初传入我国境内,随后迅速扩散蔓延。根据国家林业局公告(2019 年第 4 号),2019 年我国大陆地区松材线虫发病区已经扩张至18 个省市自治区的531 个区县,对我国松林生态系统构成严重威胁。松材线虫的生活史可分为繁殖型周期和扩散型周期[2,5]。在适宜的环境下,松材线虫进入繁殖型周期(egg、L1、L2、L3、L4和 adult),种群数量不断增加,造成松树发病死亡。在不良环境条件下,松材线虫进入扩散型周期即滞育型周期,包括扩散型三龄幼虫(DL3)和扩散型四龄幼虫(Dauer,DL4)两个阶段[6]。当松墨天牛Monochamus alternatus的蛹羽化为成虫时,扩散型三龄幼虫被诱导形成扩散型四龄幼虫,进入松墨天牛体内[7-9]。扩散型四龄幼虫口针退化不取食,依靠消耗体内能量运动[8]。天牛成虫羽化后取食健康松树时,扩散型四龄幼虫转移至健康松树,并在松树体内适宜的环境条件下解除滞育,转型并发育为成虫,进入繁殖周期开始大量繁殖危害松树[10]。因此,松材线虫扩散型四龄幼虫向成虫的转型是其完成危害松树生活史的重要环节。
松材线虫在扩散型四龄幼虫阶段完成了从病死松树进入天牛成虫以及从天牛成虫进入健康松树的传播过程[8]。目前对于松材线虫扩散型四龄幼虫的研究表明,松材线虫扩散型四龄幼虫与繁殖型幼虫相比较会发生生理和结构上的特异性变化,例如松材线虫扩散型四龄幼虫的表皮增厚、生殖腺发育停滞、脂滴增加、口针退化等以利于度过不良环境[8,11-12]。松材线虫扩散型四龄幼虫是扩散型三龄幼虫在天牛分泌的脂肪酸乙酯促进下发育形成的,并且发现松材线虫Insulin 信号通路的daf-16和 daf-12 基因上调表达参与此过程[8,13]。而扩散型四龄松材线虫能够受到天牛分泌的蛔甙(ascarosides)吸引从而进入天牛,伴随天牛取食健康松树进行传播[9]。松材线虫扩散型四龄幼虫除了可以受到天牛的吸引以外,还可以受到松树化学物质的吸引,在传播媒介天牛取食健康松树的新鲜树叶过程中转移至健康松树中[14]。松材线虫扩散型四龄幼虫在接触松树之后,如黑松Pinus thunbergii、赤松Pinus densiflora、五针松Pinus parviflora、落叶松Larix leptolepis和雪松Cedrus deodara等,均可以促进松材线虫向成虫的转型,而无松树刺激则转型不明显[6]。另外,研究表明松树挥发物单萜类α-蒎烯、β-蒎烯、3-蒈烯、β-月桂烯和柠檬烯均可以促进松材线虫扩散型四龄幼虫向成虫的转型[15]。然而,目前关于松材线虫扩散型四龄幼虫向成虫转型过程的研究尚不多见,其他树种对于松材线虫转型的影响也未见报道。
综上所述,笔者以松材线虫病防治关键龄期扩散型四龄幼虫为研究对象,通过光学显微镜观察了松材线虫DL4 转型的形态结构变化过程,以及油红染色后的脂肪特征变化,分析了松科植物对于转型过程的影响,为阐明松材线虫扩散型四龄幼虫向成虫转型的作用机理奠定基础,也为松材线虫病的科学防治提供新的思路。
1 材料与方法
1.1 扩散型四龄松材线虫收集 解剖松墨天牛获得扩散型四龄松材线虫。为了保证扩散型四龄松材线虫的虫态一致性,于4—8 月天牛羽化高峰期,在浙江和广东等省松材线虫病发生区的松林中悬挂诱捕器(杭州费洛蒙生物科技有限公司),收集松墨天牛,将松墨天牛肢解在含有无菌水的培养皿中,放置2 h。待松材线虫从松墨天牛气管中游出后,用移液器吸出移至新的无菌水培养皿中。将收集到的扩散型四龄松材线虫放入4 ℃冰箱待用。
1.2 扩散型四龄松材线虫转型过程形态观察 在直径35 mm 培养皿中,加入4 mL 无菌水,放置1 块5 mm×5 mm 的马尾松木片作为诱导物,加入100头扩散型四龄松材线虫。每隔12 h 观察1 次线虫状态,并拍照(CZX51 and BX51,Olympus,Japan)。
1.3 扩散型四龄松材线虫转型过程油红染色 采用油红对转型过程中的扩散型四龄松材线虫进行染色。配制0.01 g/mL 的油红溶液(Sigma,USA),溶剂为乙醇,配制完成后震荡混匀2 min,经过0.22 μm的过滤器过滤,保存于4 ℃冰箱待用。使用时,将配制完成的油红溶液再次过滤,室温放置10 min,取100 μL于1.5 mL 离心管中,加入等体积的上述不同转型时期的松材线虫,染色12 ~16 h 后,将线虫置于水中,挑取相应转型时期线虫观察、拍照。
1.4 多种松科植物对扩散型四龄松材线虫转型的影响 在天牛羽化高峰期选择银杏Ginkgo biloba、红豆杉Taxus chinensis、圆柏 Sabina chinensis、侧柏Platycladus orientalis、雪松 Cedrus atlantica、油松Pinus tabuliformis、黑松和马尾松 P.massoniana 的枝条,测试不同植物对于松材线虫扩散型四龄幼虫转型的影响。在35 mm 培养皿中,加入4 mL 无菌水,放置1 块5 mm ×5 mm 的植物枝条作为诱导物,对照未加任何植物枝条,加入30 头扩散型四龄松材线虫,2 d 后观察转型为成虫的数量,并计算转型率。试验重复3 次。
1.5 数据分析 采用SPSS 18.0 中的单因素方差分析对试验数据进行分析,数值为平均数±标准误。
2 结果与分析
2.1 扩散型四龄松材线虫向成虫转型过程形态
在马尾松木片诱导下,松材线虫扩散型四龄幼虫可在36 ~48 h 完成转型。整个转型过程可划分为5个阶段,包括扩散型四龄(DL4)、转型前期(TP)、转型中期(TM)、转型后期(TA)和转型末期(TT)。松材线虫DL4 阶段,头部钝圆,无溢缩,尾巴尖,无性别特征;TP 阶段,线虫呈现竖直状态,轻微摆动,可见头部蜕皮,出现口溢缩,尾部膨大;TM 阶段,蜕皮明显,线虫开始弯曲,出现性别特征,但是不显著;TA 阶段,进一步完成蜕皮,线虫成虫状态表现明显,雌虫出现阴门盖,雄虫出现交合刺;TT 阶段完成蜕皮(图1)。


图1 松材线虫扩散型四龄向成虫转型过程显微镜观察Fig.1 Transformation process of B.xylophilus DL4 observed by microscope
2.2 扩散型四龄松材线虫向成虫转型过程脂肪染色观察 DL4 阶段脂肪多且密集,染色较深,说明整体含量很高;头部颜色较浅,脂肪含量较少,腹部和尾部颜色较重,脂肪含量较高。TP 阶段脂肪含量迅速下降,仅有头部、尾部有区域化脂肪染色,但无明显颗粒感,而腹部和体壁内侧仍然存在少量的脂肪。TM 阶段,头部脂肪颗粒消失,腹部体壁内侧和尾部仍有少量脂肪存在。TA 阶段,雌虫头部脂肪颗粒消失,其他部位均有脂肪存在;而雄虫头部和尾部均无脂肪颗粒染色,仅有虫体中间部分有脂肪存在。TT 阶段与TA 阶段相似,但是TT 阶段松材线虫表现出明显的脂肪颗粒感,且头部出现较弱的个别脂肪颗粒(图2)。另外,松材线虫扩散型四龄幼虫有一部分不能完成转型,形态上依然保持DL4 阶段形态,但是体内脂肪颗粒大量减少,被消耗殆尽(图2,DL4 UT)。
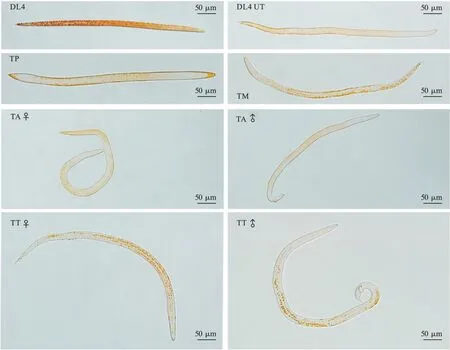
图2 松材线虫扩散型四龄向成虫转型过程脂肪染色显微镜观察Fig.2 Lipid status during transformation process of B. xylophilus DL4 observed by microscope
2.3 多种松科植物对扩散型四龄松材线虫转型的影响 非松科植物银杏对松材线虫转型没有影响,与空白对照组接近,松材线虫转型率约为13%。松科植物对松材线虫扩散型四龄幼虫转型均有显著的促进作用,红豆杉和雪松促进下松材线虫扩散型四龄幼虫的转型率约为30%,而圆柏、侧柏和油松促进转型更为明显,在其刺激下,松材线虫扩散型四龄幼虫的转型率约为55%,促进效果最为显著的为黑松和马尾松,在其刺激下,松材线虫扩散型四龄幼虫向成虫转型的转型率达到了80%以上。测试的松科植物中,雪松、油松、黑松和马尾松为已知的松材线虫寄主(图3)。
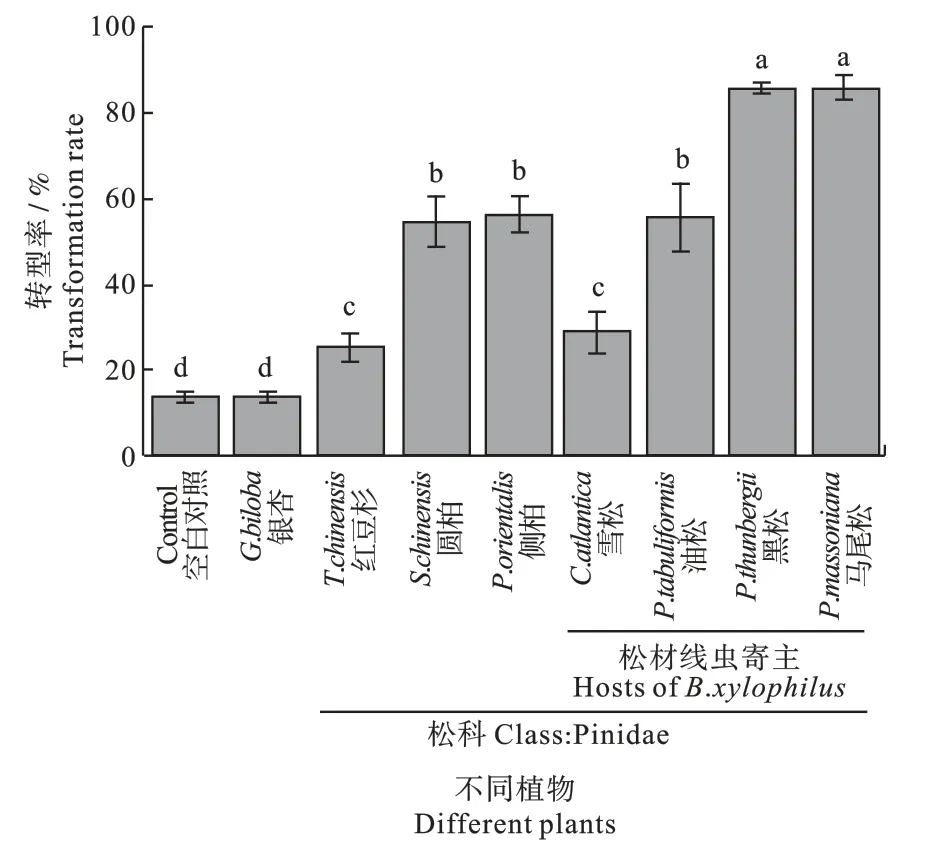
图3 多种松科植物对扩散型四龄松材线虫转型的促进作用Fig.3 Promotion of Pinidae trees to B.xylophilus DL4 transformation
3 结论与讨论
松材线虫由扩散型四龄幼虫向成虫转型是其危害松树的关键步骤,笔者观察并定义了松材线虫转型过程的5 个阶段,分别为DL4、转型前期(TP)、转型中期(TM)、转型后期(TA)和转型末期(TT)。发现转型过程中脂肪含量在DL4 阶段含量很高,但是在TP 阶段即发生了大部分的转化,而后各个阶段脂肪含量相对较少。选用的松科植物红豆杉、圆柏、侧柏、雪松、油松、黑松和马尾松均促进DL4 的转型且效果明显,其中以黑松和马尾松木块刺激下松材线虫转型率最高,而非松科植物银杏无转型促进效果。该研究为进一步探明松材线虫由扩散型四龄幼虫向成虫转型的分子及调控机制奠定基础,同时也为松材线虫的科学防治提供新的思路和理论基础。
前人仅对松材线虫扩散型四龄幼虫与成虫进行比较研究,发现松材线虫扩散型四龄幼虫的表皮增厚、生殖腺发育停滞、脂滴增加、口针退化等以利于度过不良环境,并未对线虫转型的动态过程进行深入研究[8,11-12]。笔者以松材线虫扩散型四龄幼虫为研究对象,通过光学显微镜观察了松材线虫扩散型四龄幼虫转型的形态结构变化,更加深入了解其变化过程。
与繁殖型幼虫相比,扩散型幼虫的体内脂肪含量会增加,以利于其抵抗干燥、高温或者食物短缺等不良环境[16-18]。例如秀丽隐杆线虫Caenorhabditis elegans 当环境不利时,如食物缺乏、种群密度增加、温度升高等,C.elegans 形成类似于松材线虫扩散型的dauer 幼虫[16]。该阶段C.elegans 幼虫进入滞育期,不再进食且身体结构特化,如身体辐射状缢缩、口被内部形成的栓所封闭、表皮形成特化的脊、体内脂肪粒增多等,可以抵御不良外界环境。当外界环境恢复有利条件时,如充足的食物、稀疏的种群密度和温和的温度等,dauer 幼虫会转型进入繁殖周期。松材线虫扩散型四龄幼虫与繁殖型相比较,其肠道和皮下存在大量脂滴,其中包含一些高电子密度物质[11],我们的研究也表明DL4 阶段含有大量脂肪。另外,松材线虫扩散型四龄幼虫有一部分不能完成转型,形态上依然保持DL4 阶段形态,但是体内脂肪颗粒大量减少,被消耗殆尽。这些不转型的扩散型松材线虫可能未接收到转型启动信号不能转型,但是线虫一直处于运动状态,需要不断消耗脂肪维持能量供给,因此脂肪被消耗掉。
在不同松科植物刺激条件下,松材线虫能够迅速完成转型,发育为成虫。非松科植物银杏对松材线虫转型没有影响,而松科植物对松材线虫扩散型四龄幼虫转型均有显著的促进作用,其中以黑松和马尾松促进效果最为显著。前人研究也表明松材线虫扩散型四龄幼虫在接触松树之后,黑松、赤松、五针松、落叶松和雪松等均可以促进松材线虫向成虫的转型,而无松树刺激则转型不明显[6]。另外,松树挥发物单萜类α-蒎烯、β-蒎烯、3-蒈烯、β-月桂烯和柠檬烯均可以促进松材线虫扩散型四龄幼虫向成虫的转型[15]。由此,可以推测松科植物促进松材线虫扩散性四龄幼虫转型与其含有的萜烯类物质有关。
萜烯类物质促进松材线虫扩散型四龄幼虫向成虫转型,其机理目前尚不清楚。在 C.elegans 的dauer 转型过程中,当遇到环境信号(食物、温度和密度等)刺激时,细胞膜上的G-蛋白偶联化感受体与配体化学分子结合,启动鸟苷酸环化酶途径,导致神经元细胞内环磷鸟嘌呤核苷(cGMP)浓度升高,激活cGMP-门控离子通道,将胞外信号传递进入胞内,激活 Insulin 和 TGF- β 通路的信号分子[19]。这些信号分子在内分泌细胞表面与各自受体结合,分别启动相应通路信号转导。最终两条信号通路共同调控daf-9 的表达,产生的DA 与靶组织DAF-12 结合后,促进繁殖发育,抑制dauer 形成。相反,当DA 含量下降时,空载核受体DAF-12则促进dauer 形成,抑制繁殖发育[20]。研究表明,松材线虫扩散型四龄幼虫是由扩散型三龄幼虫在天牛分泌的脂肪酸乙酯的促进下发育形成的,并且发现松材线虫Insulin 信号通路的daf-16 和daf-12 基因上调表达参与此过程[8,13]。因此,C.elegans 的dauer 形成因素及基因表达调控为进一步研究松材线虫扩散型四龄幼虫向成虫转型的机制提供了良好的理论基础,也为松材线虫的科学防治提供新的思路。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2021年16期)2021-11-26
小哥白尼(野生动物)(2021年1期)2021-07-16
西北农业学报(2020年12期)2020-12-14
新农民(2020年5期)2020-12-10
绿色中国(2019年13期)2019-11-26
中国森林病虫(2019年5期)2019-10-11
小学生必读(低年级版)(2018年10期)2019-01-04
故事作文·低年级(2018年10期)2018-10-25
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17
