
稻草不同还田量对土壤动物群落结构的影响
2020-01-18 02:16罗熳丽段均华卢昌泰肖玖金
浙江农林大学学报 2020年1期
罗熳丽, 段均华, 姚 恒, 卢昌泰, 肖玖金, 张 健
(1. 四川农业大学 生态林业研究所, 四川 成都611130; 2. 四川农业大学 林业生态工程省级重点实验室, 四川 成都 611130; 3. 四川农业大学 旅游学院, 四川 成都611830; 4. 广元市朝天区林业和园林局, 四川 广元 628012)
土壤动物及土壤微生物的活动能够改善土壤的通气状况、 养分有效性, 在土壤形成、 发育、 演化及土壤肥力形成演变中发挥着重要的作用。 土壤动物数量繁多, 通过参与一系列生命活动, 直接或间接改变土壤的性质[1], 对土壤形成、 发育及生态系统物质循环都具有重要意义。 稻草还田是一种有效的农田培肥措施[3], 在避免稻草焚烧造成环境污染的同时, 也为农业生产提供有机肥源。 作为重要肥料来源和潜在的碳库能源[2], 还田的稻草在增加土壤养分[4], 培肥地力, 改善土壤理化性状[5], 优化农田生态环境, 提高作物产量与品质[6-9]等方面意义重大。 本研究以未进行稻草还田的处理为对照, 设置不同质量稻草还田处理, 并调查不同处理下农田土壤动物群落特征, 分析不同稻草还田量对土壤动物群落结构的影响, 旨在为农田耕地环境的保护及可持续农业发展提供理论依据。
1 材料与方法
1.1 研究区概况
研究区四川省都江堰市浦阳镇金凤村(31°0′23″N, 103°37′18″E)属四川盆地亚热带湿润气候, 年平均气温为15.2 ℃, 年降水量为1 243.8 mm, 空间分布不均, 呈东南向西北减少, 无霜期269.0 d, 年蒸发量为930.1 mm。 土壤类型为山地黄壤。 样地为水稻Oryza sativa-油菜Brassica campestris轮作模式, 水稻品种为‘川优6203’ ‘Chuanyou 6203’, 油菜品种为‘绵油88’ ‘Mianyou 88’。
1.2 研究方法
2016 年4 月, 在耕地内按照具有代表性的原则设置2 个面积均为10 m × 30 m 的样地; 各样地内设置面积为5 m × 5 m 样方各4 个, 保持各样方间间距大于2 m, 其中4 个为重复样地。 随机编号, 按照单位面积稻草的平均产生量, 选取最新收获的水稻稻草, 切成3~5 段后按0 (ck)、 0.2 (T5)、 0.4 (T10)、0.8 kg·m-2(T20)的稻草均匀铺撒在样方上进行还田。
分别在还田2 个月和5 个月时, 在各样方内按 “品” 字形布点, 随机设置3 个大小为50 cm × 50 cm 的小样方, 采集0~5、 5~10、 10~15 cm 深度土层。 手捡法分层收集土壤中的大型动物, 并用体积分数为75%的乙醇将动物杀死, 带回实验室。 环刀法(r=5 cm,V=100 cm3)自上而下依次取土样, 每层各取2 个, 用尼龙网包好贴上标签迅速放入黑布袋带回实验室; 用Tullgren[10]干漏斗和Baermann[11]湿漏斗分离土壤中的中小型土壤动物, 分离时间为48 h, 其中湿生土壤动物每4 h 观察1 次。 干生分离出的土壤动物用盛有体积分数为75%乙醇的器皿收集, 湿生土壤动物用清水收集, 解剖镜下进行分类鉴定和数量统计。
所得土壤动物用双目解剖镜(Leica, EZ4HD)进行观察, 参照《中国土壤动物检索图鉴》[12]、 《昆虫分类检索》[13]、 《幼虫分类学》[14]进行分类鉴定, 一般鉴定到科, 同时统计个体数量。
1.3 数据分析与处理
土壤动物多样性分析: 采用Shannon-Wiener 多样性指数(H′)、 Margalef 丰富度指数(D)[15]、 Pielou均匀度指数(J)[16]、 Simpson 优势度指数(C)[17]。 计算公式如下: Shannon-Wiener 多样性指数式中:Pi=ni/N,ni为第i个类群的个体数;N为所有类群的个体数。 Margalef 丰富度指数D=(S-1)/lnN。式中:S为类群数,N为所有类群的个体数。 Pielou 均匀度指数J=H′/lnS。 式中:H′ 为Shannon-Wiener多样性指数,S为类群数。 Simpson 优势度指数其中:ni为第i个类群的个体数;N为所有类群的个体数。
类群数量等级划分: 个体数量大于捕获总量的10.0%者为优势类群(+++), 大于等于1.0%小于等于10.0%者为常见类群(++), 小于1.0%者为稀有类群(+)。
采用Excel 2010 和SPSS 22.0 完成数据处理和分析。 用单因素方差分析(one-way ANOVA)对不同样方间土壤动物群落组成进行检验; 用LSD(方差齐性)法进行多重比较, 显著性水平设定为P=0.05[18]。
2 结果与分析
2.1 土壤动物种类和数量组成
本研究共捕获土壤动物1 194 只, 隶属于3 门11 纲19 目44 个类群, 类群数排序为T20>T10>T5>对照。 优势类群为线虫纲Nematoda 和懒甲螨科Nothridae, 分别占总捕获量的72.16%和11.09%; 常见类群为蚁科Formicidae、 线蚓科Enchytraeidae、 绥螨科Sejidae、 丽甲螨科Liacaridae、 双翅目Diptera 幼虫, 分别占总量的4.33%、 4.00%、 2.41%、 1.95%和1.59%; 棘科Onychiuridae、 跳虫科Poduridae、露尾甲科Nitidulidae 等构成稀有类群, 占总量的2.47%。
由表1 可见: 还田2 个月后各样地的优势类群都为线虫纲和懒甲螨科, 其中: T20样地2 次捕获的土壤动物包含30 个类群, 平均密度4.13×104只·m-2; T10样地2 次捕获的土壤动物隶属于20 科, 平均密度为4.83×104只·m-2; T5样地2 次捕获的土壤动物隶属于21 科, 平均密度4.73×104只·m-2; 对照样地2 次捕获的土壤动物隶属于11 科, 平均密度3.70×104只·m-2。 还田5 个月后T20、 T10、 T5样地的优势类群都为线虫纲, 对照样地优势类群为线虫纲和蚁科, 其中: T20样地2 次捕获的土壤动物包含22 科,优势类群为线虫纲, 平均密度9.32×104只·m-2; T10样地2 次捕获的土壤动物隶属于20 科, 平均密度8.48×104只·m-2; T5样地2 次捕获的土壤动物隶属于19 科, 平均密度4.75×104只·m-2; 对照样地2 次捕获的土壤动物隶属于11 科, 平均密度4.60×104只·m-2。
2.2 土壤动物群落水平分布特征
由图1A 可知: 还田5 个月后各样地的土壤动物密度均高于还田2 个月后的土壤密度。 还田2 个月后, 各样地的平均密度排序为T10>T5>T20>对照, 各样地间差异不显著(F=0.105,P=0.956)。 还田5 个月后, 各样地的平均密度排序为T20>T10>T5>对照, 样地间差异极显著(F=6.702,P=0.003); 其中, T20样地与对照、 T5样地的平均密度均呈极显著差异(P<0.01), T10与T5样地的平均密度均呈极显著差异(P<0.01), 与对照样地呈显著差异(P<0.05)。
T20土壤动物的类群数均高于T10、 T5和对照样地的土壤动物。 统计分析显示(图1B): 还田2 个月后样地间土壤动物类群数差异性极显著(F=9.312,P=0.001), 其中, T20样地和T10、 T5、 对照样地分别呈极显著差异(P<0.01); 还田5 个月后T10和T20样地的土壤动物类群数呈显著差异(F=1.299,P=0.011)。
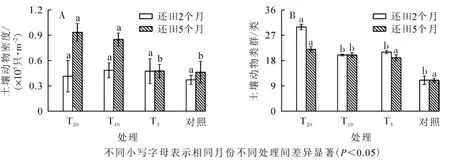
图1 各生境土壤动物水平分布变化Figure 1 Each horizontal distribution of soil fauna habitat change
2.3 不同体型土壤动物群落的水平分布特征
按照尹文英[12]对土壤动物体型的划分, 土壤动物可分为大型和中小型2 类。 由图2 可以看出: 与对照相比, T20、 T10、 T5样地大型、 中小型土壤动物的平均密度均较高。 还田2 个月后, 各处理大型土壤动物平均密度排序为T20>T10>T5>对照, 其中T20样地和对照差异显著(P<0.05), 其他样地间差异不显著; 中小型土壤动物以T10样地平均密度最高, 对照样地最低, 差异均不显著。 还田5 个月后, T20样地大型土壤动物密度最高, 占总密度的41.44%, 其余依次为T5(35.14%)、 T10(18.02%)、 对照(5.40%)。统计分析显示: 还田5 个月后, 各样地中小型土壤动物平均密度差异极显著(F=6.759,P=0.003), 对照分别与T10、 T20呈显著差异(P<0.05), T5分别与T10、 T20呈显著差异(P<0.05)。
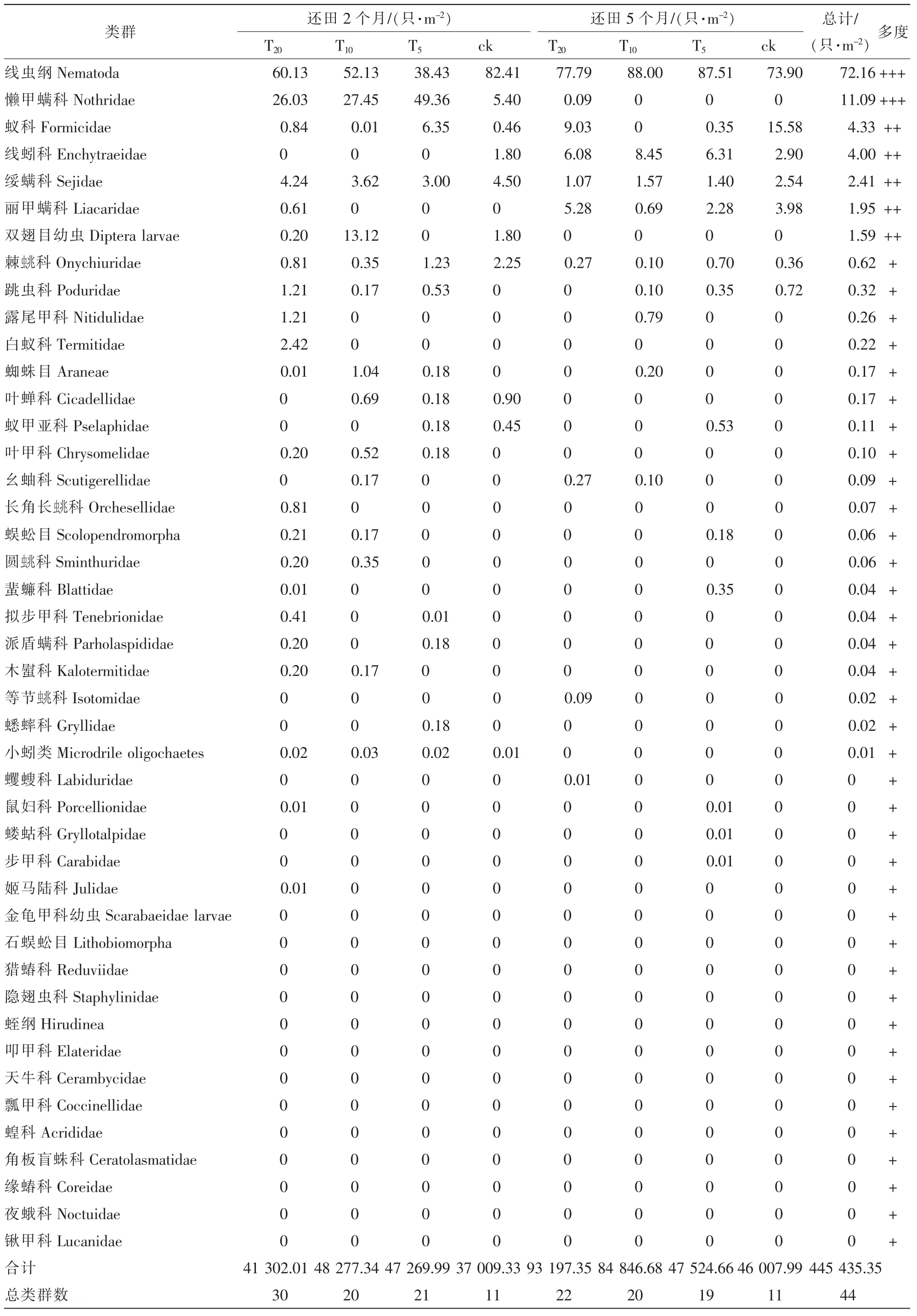
表1 不同样地土壤动物群落密度统计Table 1 Compositions of soil fauna community in the different plots
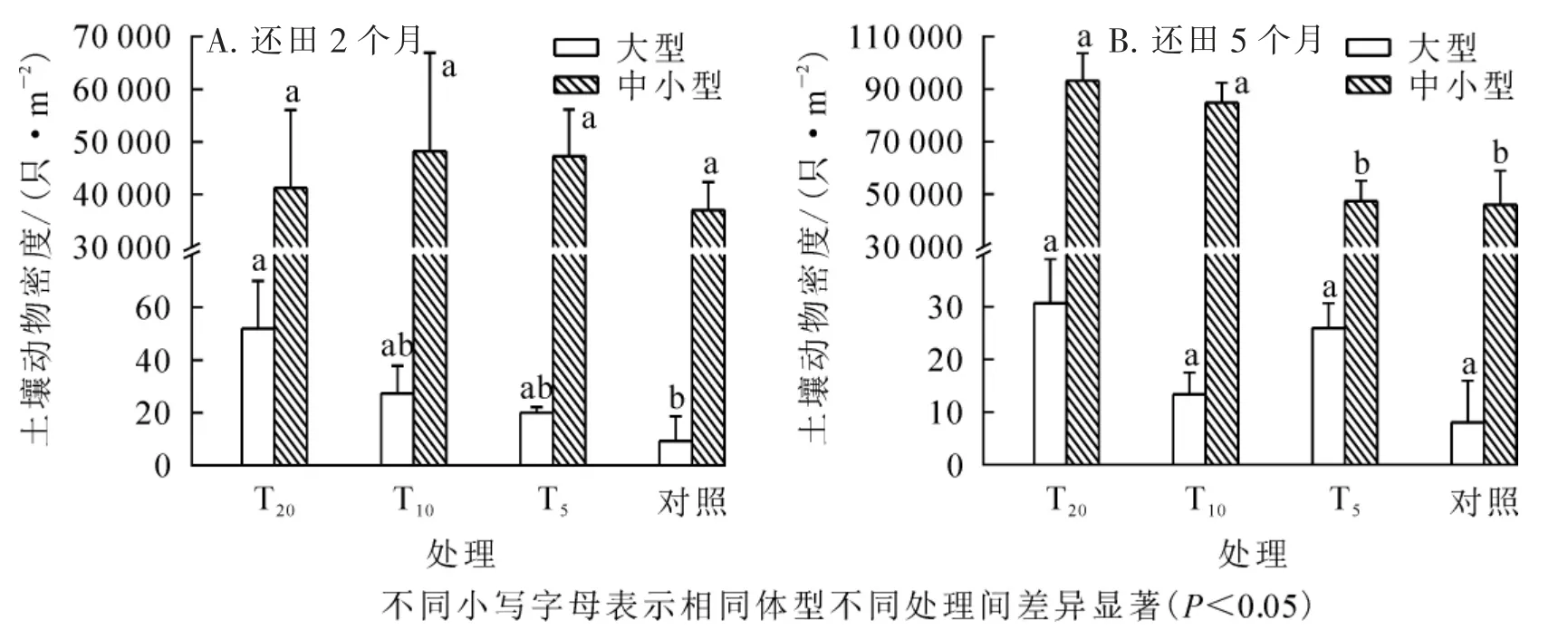
图2 各生境不同体型土壤动物分布变化Figure 2 Each horizontal size distribution of soil fauna in the different levels of change
2.4 土壤动物群落的垂直分布特征
对不同稻草还田量下0~5、 5~10 和10~15 cm 层的土壤动物密度进行比较, 结果显示: 0~5 cm 层土壤动物平均密度最高, 随着土层加深土壤动物密度降低, 具有明显的表聚性(图3A)。
方差分析显示(图3B): 还田2 个月后各样地所有土层土壤动物平均密度差异均不显著(P>0.05);还田5 个月后各样地10~15 cm 土层土壤动物平均密度差异不显著(F=1.556,P=0.237), 0~5 cm 层(F=6.099,P=0.005)、 5~10 cm 层(F=5.942,P=0.006)差异显著。 0~5 cm 土层中, 对照与T20样地土壤动物密度差异显著(P<0.05), T5与T10、 T20样地差异显著(P<0.05)。
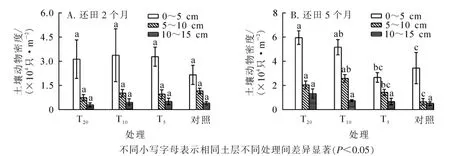
图3 各生境土壤动物垂直分布变化Figure 3 Each vertical distribution of soil fauna habitat change
2.5 土壤动物的多样性特征
稻草还田提高了土壤动物的多样性指数。 还田2 个月后各处理下土壤动物多样性指数(H′)、 丰富度指数(D)均高于对照, 优势度指数(C)均低于对照, 均匀度指数T20和T5高于对照, 但T10低于对照。 还田5 个月后对照土壤动物多样性指数和均匀度指数最高, T10处理下土壤动物优势度指数最高, T20处理下土壤动物丰富度指数最高(表2)。
方差分析结果显示: 还田2 个月后, 各样地中土壤动物优势度指数(F=1.810,P=0.184)、 多样性指数(F=2.049,P=0.145)、 均匀性指数(F=0.270,P=0.846)的差异均不显著, 丰富度指数(F=9.764,P=0.001)T20样地与T10、 T5、 对照呈极显著差异(P<0.01)。 还田5 个月后, 各样地中土壤动物优势度指数(F=1.436,P=0.267)、 多样性指数(F=1.795,P=0.186)差异均不显著, T20与T10样地的丰富度指数差异显著(P<0.05), 对照与T10样地的均匀度指数差异显著(P<0.05)。
3 讨论
目前, 稻草还田已经作为农业上培肥地力的一项技术而普遍利用。 本研究发现: 还田2 个月和5 个月后, 土壤动物类群数排序均为T20>T10>T5>对照, 土壤动物密度排序为T10>T5>T20>对照。 整体来看, 土壤动物平均密度和类群数差异不显著, 原因可能有以下几点。 首先, 实验在夏季进行, 土壤温度高、 湿度大, 微生物活动旺盛, 分解快, 还田5 个月后(9 月)未发现明显稻草的分解物, 说明腐熟高峰已过, 土壤营养处于下降状态, 因此土壤动物个体数量和类群数降低。 其次, 稻草覆盖的还田方式增加了农田表层腐殖质的土壤水分和有机质, 提高了土壤保水保墒能力, 喜湿土壤动物幼虫数量明显增加,土壤动物类群数量也随之增加, 其中, 稻草还田处理的土壤线虫数量最高达到ck 处理的2.5 倍; 再次,单位面积上稻草还田较多, 则分解后养分较多, 有利于创造适宜土壤动物生存繁衍的环境[19-21]。 相比于稻草还田处理, 对照土壤贫瘠, 土壤肥力较差, 因此土壤动物类群数和土壤动物密度都最低[22]。 这与在楠木人工林凋落物和草本层对土壤动物群落的影响中得到相似的结论[23]。
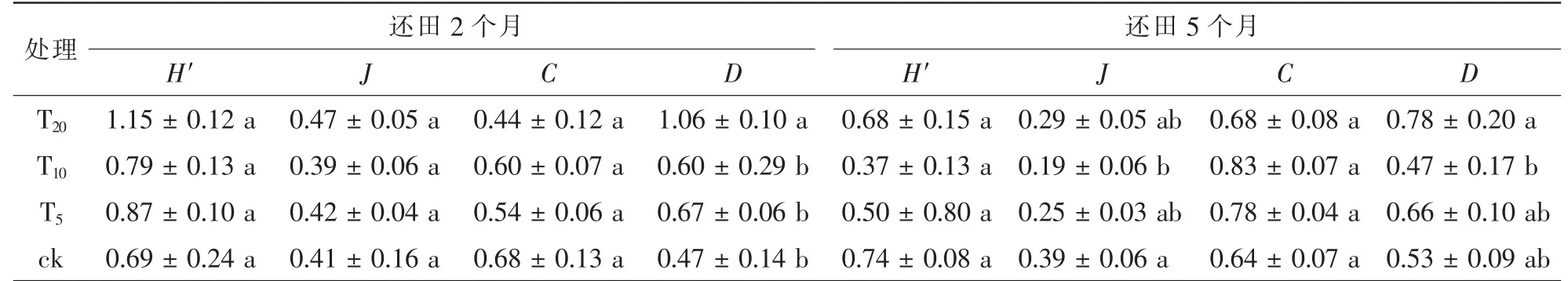
表2 稻草不同还田量下土壤动物群落的多样性特征Table 2 Diversity characteristics of soil fauna in each habitat
各样地土壤动物密度剖面及类群分布具明显的表聚性特征, 2 次采样所采集到的大型、 中小型土壤动物的数量有一定的差异, 随着季节的不同土壤动物的多样性也各不相同。 还田2 个月和还田5 个月,不同量稻草还田对土壤动物多样性的影响不同, 具体来说, 土壤动物数量与类群在不同还田量稻草处理下差异明显, 稻草还田量大的样地土壤动物数量与类群明显多于稻草还田量少的样地。 稻草还田2 个月后各样地上土壤动物多样性指数、 均匀度指数和丰富度指数均高于还田5 个月的样地, 以线虫纲密度增加最为明显, 可能是因为稻草还田后线虫等中小型土壤动物对土壤环境敏感性强、 响应明显; 优势度指数则低于还田5 个月后, 与此同时对照组不同月份各指数差异变化不大, 说明稻草还田后, 样地的土壤动物种群更加丰富, 土壤的生态环境相对稳定良好, 一段时间后优势种群开始突出。
综上所述, 与未进行稻草还田处理的普通样地相比, 稻草还田显著增加了土壤动物密度和类群数,影响了区域内土壤动物群落结构; 稻草还田通过改变土壤理化性质, 促进了土壤动物的多样性和生态系统的稳定性, 提高了作物质量和产量[22,24]。 基于对生态系统稳定性及生产效率等因素的考虑, 建议农业生产上稻草切碎后覆盖还田选择0.8 kg·m-2还田量。 本次研究历时较短, 仅研究了稻草覆盖还田对土壤动物群落的影响, 今后应继续对稻草还田腐烂程度等影响因素的土壤动物动态变化特征进行监测研究,为农业生产提供更加科学的依据。
猜你喜欢
今日农业(2022年14期)2022-09-15
经济林研究(2022年2期)2022-07-20
贵州畜牧兽医(2022年3期)2022-06-28
农药科学与管理(2021年11期)2022-01-12
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
农业科技与信息(2020年14期)2020-12-18
西北农业学报(2020年12期)2020-12-14
载人航天(2020年3期)2020-07-02
绿色科技(2019年14期)2019-11-19
