
缺血后处理对全脑缺血再灌注大鼠海马区神经元自噬的影响
2020-01-10 00:35:32刘瑶韩影许丽雅赵雅宁刘宇刘俊杰李建民
中国老年学杂志 2020年1期
刘瑶 韩影 许丽雅 赵雅宁 刘宇 刘俊杰 李建民
(华北理工大学,河北 唐山 063000)
脑缺血已经成为严重危害中老年人群生命安全和生活质量的严重疾病,由于大脑对氧的需求量比较大,比其他器官更容易产生再灌注损伤〔1〕,所以研究怎样减轻缺血再灌注损伤对治疗脑缺血具有十分重要的意义。缺血后处理是指在缺血后再灌注早期对缺血组织进行反复、短暂性缺血再灌注处理,使组织对缺血产生耐受性,从而减轻再灌注损伤,经研究发现缺血后处理可以减少梗死面积、保护内皮细胞功能,是缺血再灌注损伤较有力的内源性保护机制,在临床上缺血性心、脑及外周血管疾病的防治具有良好的应用前景。近年来自噬在神经系统疾病中的作用及其作用机制受到国内外学者广泛关注。靳奥洁等〔2〕通过对大鼠脑缺血再灌注模型研究,指出自噬参与脑缺血再灌注损伤的过程。适度自噬可以发挥神经保护作用,而过度自噬引起自噬性细胞死亡。脑缺血后处理作为一种新型的脑保护机制,其明显减轻了脑缺血再灌注损伤,经研究发现缺血后处理对神经细胞的保护作用可能与蛋白激酶途径、促分裂原活化蛋白激酶(MAPK)途径、炎症等有关〔3~5〕,但其与自噬之间的联系目前尚不清楚,故本文通过检测自噬相关蛋白Beclin-1、LC3-Ⅱ 的表达变化,探讨脑缺血后处理的脑保护作用机制。
1 材料与方法
1.1动物 成年健康雄性SD大鼠72只,10~12周龄,体重(300±50)g,来自华北理工大学实验动物中心,合格证号SCXK(京)2009-003;清洁级,自由饮食,室温饲养。
1.2主要试剂与仪器 兔抗Beclin1多克隆抗体、兔抗LC3多克隆抗体、兔抗β-actin多克隆抗体由北京博奥森生物技术有限公司提供。免疫组化试剂盒购自北京中杉金桥生物技术有限公司。Olympus光学显微镜由日本Olympus公司提供。Nikon摄影生物光学显微镜和Bio-Rad凝胶成像系统为日本株式会社尼康产品。
1.3方法
1.3.1动物分组和造模 将72只大鼠采用随机数字法分为假手术组(Sham组)、脑缺血再灌注模型组(CIR组)、脑缺血后处理组(CIP组),每组24只。采用改良的Pulsinelli四血管闭塞法〔6〕建立大鼠全脑缺血模型。术后24 h后用无创动脉夹同时夹闭双侧颈总动脉15 min,大鼠全脑缺血成功判断标准为:大鼠昏迷、翻正反射消失、双侧瞳孔放大、眼睛颜色发白。15 min后大鼠恢复再灌注,放回笼中单独饲养直至相应时间点留取CIR组大鼠标本。CIP组大鼠首先同CIR组大鼠一样采用改良的Pulsinelli四血管闭塞法制作大鼠全脑缺血模型,然后在完全恢复再灌注前给予短暂的再灌注/缺血处理,时间各15 s,重复3次,于恢复再灌注的相应用时间点处死大鼠留取本实验大鼠脑组织标本。
1.3.2苏木素-伊红(HE)染色 在预定时间点对大鼠进行麻醉,开胸,暴露心脏,穿刺主动脉后,注射4%的多聚甲醛250~300 ml,断头取脑,于4%多聚甲醛固定脑组织中。常规包埋蜡块。连续冠状切片,片厚约为4 μm,常规脱蜡至水,经HE染色。光学显微镜高倍镜(×400)下随机观察各时间点每组均随机数字表法选取的切片,采用Motic6.0 数码医学图像采集分析系统计数每个视野的存活细胞,取其平均值进行统计分析。
1.3.3免疫组织化学方法 采用SP法。常规脱蜡至水,3%过氧化氢中浸泡,高压修复抗原;滴加一抗Beclin-1、LC3-Ⅱ(1∶200),湿盒中过夜;磷酸盐缓冲液(PBS)冲洗后滴加二抗PV6001,放于37℃温箱孵育;二氨基联苯胺(DAB)溶液显色,梯度酒精脱水,透明封片。Beclin-1、LC3-Ⅱ阳性细胞计数:光学显微镜(40×10倍)下观察脑组织切片,Olympus摄影显微镜计数海马区不重叠视野的阳性细胞,取其均值进行统计学分析。
1.3.4Western 印迹 无须进行灌注,直接于冰上断头取脑,剥离出内部双侧似月牙形状的海马,放置于EP管中,标记好后放于-80℃冰箱保存。取出海马组织,用组织裂解液提蛋白,配置标准蛋白,酶标仪绘制标准曲线,计算蛋白样品浓度,制胶,进行SDS-PAGE,转膜,洗膜,加入兔抗Beclin-1多克隆一抗(1∶800稀释)、兔抗LC3多克隆一抗(1∶800稀释),4℃冰箱过夜。TBST洗膜,加入山羊抗兔二抗(1∶2 000稀释),采用Bio-Rad凝胶成像分析系统对条带进行曝光、显影和分析。将目的蛋白与内参β-actin的光密度值进行比较,对蛋白表达含量进行半定量分析。
1.4统计学分析 采用SPSS19.0统计软件进行单因素方差分析。
2 结 果
2.1各组海马区HE染色结果 Sham组大鼠海马区神经元结构和形态正常,排列整齐紧密,未见坏死变形的细胞;与Sham组比较,CIR组大鼠海马区神经元结构损伤,各时间点存活神经元数量较明显减少(P<0.05);与CIR组比较,CIP组大鼠海马区神经元结构损伤减轻,各时间点存活神经元数量明显增多(P<0.05)。见表1,图1。
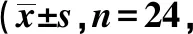
表1 各组海马区存活神经元数量个/高倍视野)
与Sham组比较:1)P<0.05;与CIR组比较:2)P<0.05

图1 镜下观察各组海马区神经元结构(HE,×400)
2.2各组海马区免疫组化结果
2.2.1各组海马区LC3-Ⅱ表达结果 Sham组大鼠海马区各个时间点可见少量LC3-Ⅱ阳性弱表达;与Sham组比较, CIR组、CIP组大鼠海马区各时间点LC3-Ⅱ表达显著增加(P<0.05),其中6 h开始增加,24 h达高峰,48 h、72 h下降;与CIR组比较,CIP组大鼠海马区各时间点LC3-Ⅱ表达显著增加(P<0.05);CIP组大鼠海马区LC3-Ⅱ表达24 h达高峰,组内各时间点比较差异有统计学意义(P<0.05)。见表2,图2。
表2 各组海马区LC3-Ⅱ阳性细胞表达个/高倍视野)
与Sham组比较:1)P<0.05;与CIR组比较:2)P<0.05;与CIP组内24 h比较:3)P<0.05,下表同
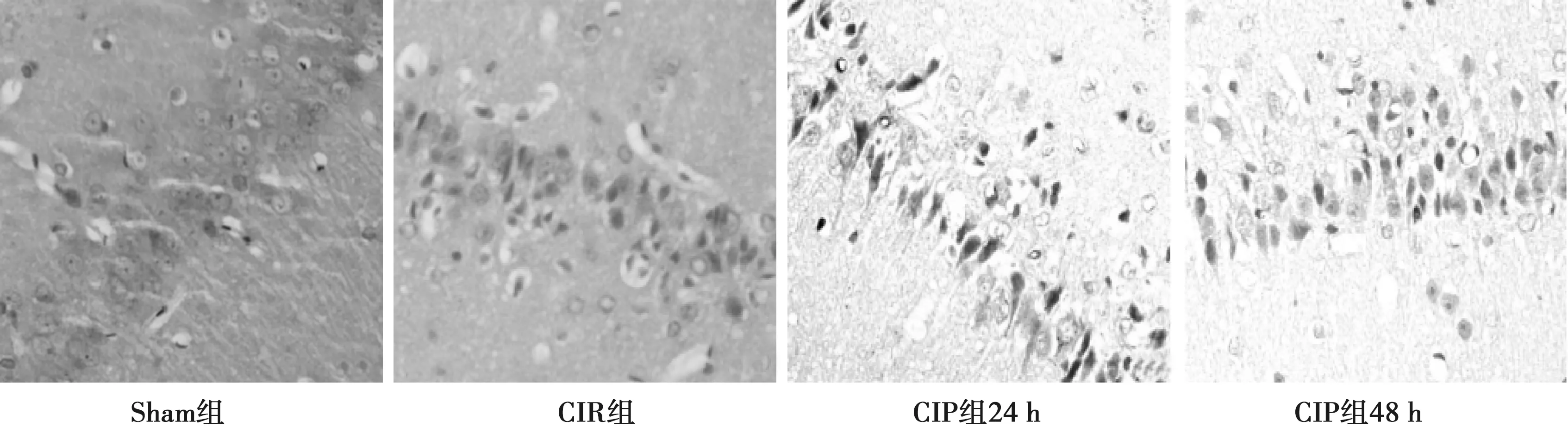
图2 各组海马区LC3-Ⅱ表达(DAB,×400)
2.2.2各组海马区Beclin-1表达结果 Sham组大鼠海马区各个时间点可见少量Beclin-1阳性弱表达;与Sham组比较,CIR组大鼠海马区各时间点Beclin-1表达显著增加(P<0.05);与CIR组比较,CIP组大鼠海马区各时间点Beclin-1表达显著增加(P<0.05);CIP组大鼠海马区Beclin-1表达24 h达高峰,组内各时间点比较差异有统计学意义(P<0.05)。见表3,图3。
表3 各组海马区Beclin-1阳性细胞表达个/高倍视野)

图3 各组海马区Beclin-1表达(DAB,×400)
2.3各组海马区Western 印迹结果
2.3.1各组海马区LC3-Ⅱ蛋白表达 Sham组大鼠海马区各个时间点见少量LC3-Ⅱ蛋白表达;与Sham组比较,CIR组大鼠海马区各时间点LC3-Ⅱ蛋白表达量显著增加,差异有统计学意义(P<0.05);与CIR组比较,CIP组大鼠海马区各时间点LC3-Ⅱ蛋白表达量显著增加,72 h表达最低,差异均有统计学意义(P<0.05)。见表4,图4。
表4 各组海马区LC3-Ⅱ蛋白Western印迹结果个/高倍视野)
与Sham组比较:1)P<0.05;与CIR组比较:2)P<0.05;与本组24 h比较:3)P<0.05;与本组72 h比较:4)P<0.05,下表同

图4 LC3-Ⅱ蛋白Western印迹结果
2.3.2各组海马区Beclin-1蛋白表达 以Beclin-1光密度值/β-actin光密度值表示Beclin-1蛋白的相对表达量。Sham组大鼠海马区各个时间点见少量Beclin-1蛋白表达;与Sham组比较,CIR组大鼠海马区各时间点Beclin-1蛋白表达量显著增加,差异有统计学意义(P<0.05);与CIR组比较,CIP组大鼠海马区各时间点Beclin-1蛋白表达量显著增加,24 h表达最高,72 h表达最低,差异均有统计学意义(P<0.05)。见图5,表5。

图5 Beclin-1蛋白Western印迹结果
表5 各组海马区Beclin-1蛋白Western印迹结果个/高倍视野)
3 讨 论
脑缺血是一种多因素的异质性疾病,死亡率和致残率都很高,影响着各个年龄组、种族和国家的人口,尤其是对老年人的损害比较严重。缺血后处理通常是指长时间严重缺血损伤后对大脑或远端肢体的血液供应进行的一次或一系列短暂的干扰,调动机体内源性物质,为脑缺血诱导的神经元丧失提供脑保护〔7〕。经研究发现,缺血后处理在减轻心肌损伤的缺血再灌注损伤,肺移植再灌注损伤,肾脏缺血再灌注损伤等发挥着重要作用〔8~10〕。2006年Zhao等〔11〕发现缺血后处理在发生脑卒中后具有促进神经功能恢复,保护脑功能的作用。通过大鼠局灶性脑缺血模型研究证实,缺血后处理能抑制细胞凋亡发挥神经保护作用〔12〕。缺血后处理通过影响缺血后再灌注和脑血流量,改善脑缺血后神经血管单位中内皮细胞、周细胞和星形胶质细胞之间的相互作用,下调协同的有害过程,包括兴奋毒性、线粒体紊乱、氧化应激和炎症进而影响了其下游的细胞信号〔13〕。本研究证实缺血后处理能够减轻脑缺血再灌注损伤。这与以往的研究一致,导致缺血后处理脑保护作用的潜在机制是复杂的,并且仍然没有得到很好解释。
自噬是一种溶酶体介导的降解过程,它清除错误折叠的蛋白质和过量或受损的细胞器,在细胞内环境的稳态中起着至关重要的作用。细胞通过溶酶体途径降解衰亡的细胞器及损伤的蛋白即是自噬〔14〕。自噬具有双重作用,当自噬适度激活时,可加速胞内蛋白质降解,一些错误折叠的蛋白质和受损的细胞器可以被降解为代谢元素并循环利用以维持细胞内稳态,抵抗能量不足,促进细胞存活;当自噬过度激活时,可导致细胞裂解,引起细胞死亡。王国峰等〔15〕通过在大鼠脑缺血前注射自噬诱导剂雷帕霉素发现细胞凋亡减少,表明自噬能减轻缺氧缺血损伤,具有神经保护作用。Beclin-1和LC3都是重要的自噬相关蛋白,被用作自噬体的可靠生物标记物,Beclin-1是酵母自噬基因Atg6/Vps30的同源基因,Beclin1表达的上调对自噬过程也是必要的,因其表达的强度与自噬的活性程度密切相关,相反,Beclin1的沉默和LC3-Ⅰ向LC3-Ⅱ转化的减少将有助于阻止自噬体的加工〔16〕。已有研究发现通过使用自噬抑制剂3-MA和渥太青霉素下调缺血再灌注后神经元中Beclin-1表达,加重了神经元坏死,而应用自噬激活剂雷帕霉素可以上调Beclin-1表达,同时发现神经元损伤明显减少,证明了自噬在缺血再灌注后发挥重要保护作用〔16,17〕。几乎在各种组织细胞均可以见到自噬现象,自噬可以通过调节受损细胞质和线粒体的降解来维持细胞内稳态,并确保细胞在应激条件下的生存,自噬在缺血性脑再灌注损伤中受多重因素调控。在本实验中,缺血后处理通过上调缺血再灌注自噬相关蛋白 Beclin-1、LC3-Ⅱ的表达,以减少缺血再灌注损伤神经元凋亡,减轻神经功能缺损症状,从而发挥其脑保护作用,且在出缺血后处理24 h时,自噬相关蛋白 Beclin-1、LC3-Ⅱ的表达明显增多。缺血再灌注损失导致过多的ROS,而ROS不能被内源性防御系统迅速清除,以往的研究发现自噬的产生与低氧诱导因子-1α、氧自由基的适度产生等有关〔18,19〕,其主要机制与增强内源性抗氧化防御系统有关。缺血后处理可诱导低氧诱导因子-1α表达上调,避免氧自由基的过度产生而导致的自噬性死亡,从而使自噬在24 h保持一个较高的水平,发挥了保护作用。
综上所述,脑缺血再灌注损伤会造成明显的神经元损伤,缺血后处理通过激活自噬减少了神经元的凋亡,减轻了脑缺血再灌注损伤。总之,随着研究的进一步进行,缺血后处理的脑保护作用已经明确,但由于其机制广泛而复杂,还未明确。而近年来自噬在心脑血管疾病中的作用越来越受到人们的关注,关于缺血后处理与自噬的关系值得探索发现,为临床脑缺血再灌注损伤的治疗提供新思路。
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05 01:33:06
今日农业(2020年20期)2020-12-15 15:53:19
创新作文(小学版)(2019年10期)2019-09-25 08:12:28
能源(2018年10期)2018-12-08 08:02:48
小学生学习指导(低年级)(2017年5期)2017-05-04 04:14:38
能源(2016年10期)2016-02-28 11:33:30
中国康复理论与实践(2015年10期)2015-12-24 05:42:43
吉林大学学报(医学版)(2015年5期)2015-12-16 15:43:56
中国体外循环杂志(2015年3期)2015-12-08 05:13:01
作文与考试·小学高年级版(2015年17期)2015-05-30 10:48:04
