
秋英属两种植物营养器官解剖结构的比较研究
2020-01-03 03:43赵智艳刘驹王菲房琦
信阳农林学院学报 2019年4期
赵智艳,刘驹,王菲,房琦
(安徽新华学院 土木与环境工程学院,安徽 合肥 230088)
秋英属(Cosmos)隶属于菊科(Compositae)管状花亚科(Carduoideae)向日葵族(Heliantheae)。该属植物约25种,分布于美洲热带。我国常见栽培的有2种,即波斯菊(Cosmosbipinnata)和硫华菊(Cosmossulphureus)[1]。这两种植物因其花期长,花色鲜艳,耐粗放管理而备受园林绿化者的青睐,常用作地被植物大面积栽植,也可用于花境、花丛或建筑的基础种植等[2]。目前有关波斯菊和硫华菊的研究主要集中在种子萌发[3]、核型分析[4]、栽培管理[5]等方面,有关其显微结构研究尚未见报道。本研究从形态解剖方面对这两种植物的营养器官进行系统研究,对其生态学特性也作了探讨,旨为秋英属植物的研究或利用提供解剖学方面的依据。
1 材料与方法
1.1 实验材料
波斯菊和硫华菊均为普通品种,其中波斯菊为矮杆,淡紫色品种,硫华菊为“金鸟”系列,花橙色。两种试材于2019年3月20日同时播种于安徽新华学院校内苗圃基地,均种植于向阳且排水良好地段。于2019年5月12日采集观察,此时二者均为始花期,选取植株根部均已进行次生生长。
1.2 实验方法
采用徒手切片法对波斯菊和硫华菊的根、茎、叶进行切片,并制成临时装片;叶片表皮及硫华菊茎部表皮毛的观察采用表皮撕取法,使用光学显微镜观测,并采用显微互动系统拍照记录。
2 结果与分析
2.1 根的解剖结构
秋英属植物的根为直根系,主根圆锥形。本研究选取波斯菊和硫华菊的主根进行显微结构观察。结果表明:二者根横切面均为近圆形,由周皮、韧皮部、维管形成层、木质部及髓构成。
周皮是能代替表皮对根起保护作用的一种次生保护组织[6]。波斯菊根的周皮外侧细胞较小,排列致密,随着根的增粗,最外层细胞有脱落现象。周皮内侧细胞与韧皮部界限不明显,二者由排列疏松、体积较大的薄壁细胞构成,细胞间隙较大。约占根横切面直径的1/4。木质部发达,约占根直径的1/2,由木薄壁细胞、导管、木纤维和木射线构成。导管近圆形,大小不一,单个、聚生或放射状排列。木射线明显。髓部明显,为菱形,由体积较大的薄壁细胞构成,约占根直径的1/8。
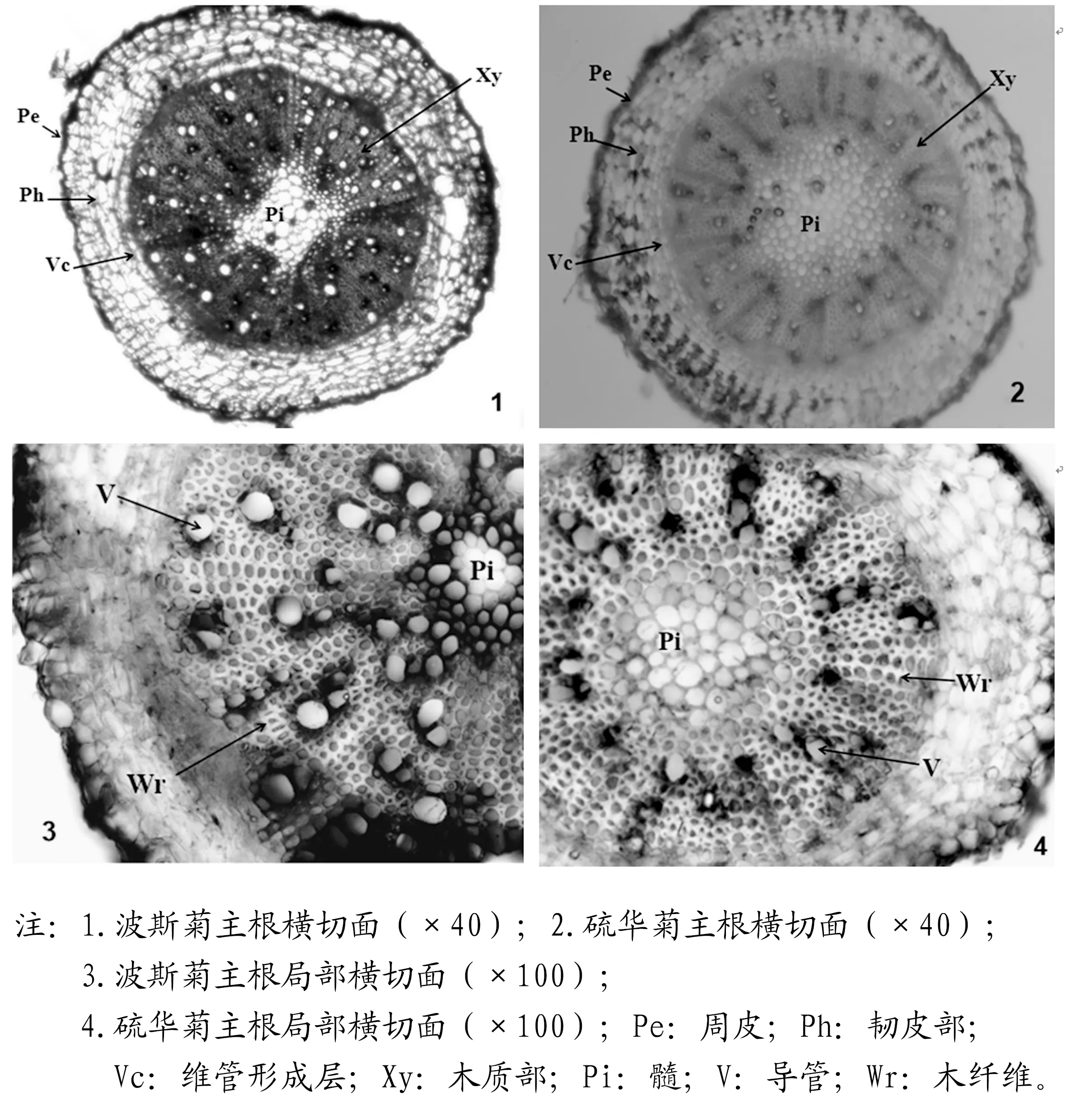
图1波斯菊与硫华菊根的显微结构
硫华菊根横切结构与波斯菊类似。不同之处为:硫华菊木质部约占根直径的1/3;导管近圆形,除少数单个、聚生排列外,大部分单列放射状排列;髓部较大,为类圆形,约占根直径1/4。
2.2 茎的解剖结构
波斯菊和硫华菊均具有典型的双子叶草本植物茎的结构。在茎的横切面上,可从外到内依次分为表皮、皮层和维管柱。表皮为一层排列整齐紧密的薄壁细胞构成。整个皮层在茎的横切面上所占比例较小,其中近表皮的3~4层外侧皮层细胞为厚角组织,可加强茎的机械支持功能,其余皮层细胞为薄壁组织。维管柱由维管束、髓和髓射线组成(图2-2、5)。维管束呈环状排列,由形成层将其串连。维管束由外到内依次为初生韧皮部、束内形成层和初生木质部。髓位于茎中央,由体积较大的薄壁细胞构成,占整个茎横切面的1/2以上。髓射线较宽,位于维管束之间,主要起径向输导作用(图2-1、2、4、5)。
两种植物茎的解剖结构相似,但也存在差异:波斯菊的茎横切面近圆形,而硫华菊的茎具棱,横切面呈齿凸状(图2-1、4);波斯菊的茎肉眼看较光滑,但显微镜下可观察到由2~3个细胞构成的细小的表皮毛,长约45μm,硫华菊的茎肉眼可看到较粗壮的表皮毛,但茎横面常观察不到表皮毛,通过撕取茎表皮,可观察到14~20个细胞构成的表皮毛,长度可达1260μm(图2-3、6);波斯菊茎横切面具18~21个维管束,硫华菊具14~16个维管束。
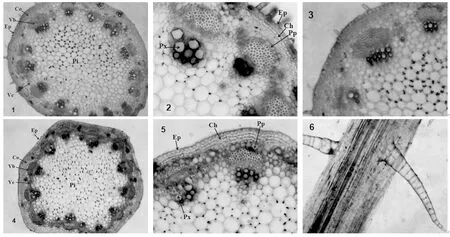
注:1.波斯菊茎横切面(×40);2.波斯菊茎局部横切面(×100);3.波斯菊茎表皮毛(×40);4.硫华菊茎横切面(×40);5.硫华菊茎局部横切面(×100);6.硫华菊茎表皮毛(×40);Ep:表皮;Co:皮层;Pp:初生韧皮部;Px:初生木质部;Pi:髓;Vb:维管束;Vc:维管形成层;Ch:厚角组织
2.3 叶的解剖结构

图3波斯菊与硫华菊叶表皮显微结构
2.3.1 叶片表皮结构 从横切面来看,波斯菊与硫华菊的叶片均由表皮、叶脉和叶肉组成。表皮由单层细胞构成,细胞排列紧密,外切向壁略向外突,均具表皮毛和角质层。
两种植物上表皮细胞形状相似,均为不规则多边形;下表皮细胞形状差异较大,其中波斯菊下表皮细胞为不规则长柱状,硫华菊表皮细胞为不规则多角棱状。
两种植物上下表皮均有气孔分布,气孔均无副保卫细胞。通过测量可知,二者气孔大小相似,且下表皮气孔大小及密度均大于上表皮气孔。硫华菊上下表皮气孔密度较波斯菊大(图3,表1)。
2.3.2 叶肉及叶脉结构 叶肉组织是植物进行光合作用的主要部分,根据叶肉组织的分化程度,可将叶片分为等面叶和异面叶。通过显微镜观察,两种植物均为异面叶,叶肉靠近上表皮为栅栏组织,靠近下表皮为海绵组织。

注:1.波斯菊叶片横切面(×40);2.波斯菊叶片局部横切面(×100);3.波斯菊叶柄横切面(×40);4.硫华菊叶片横切面(×40);5.硫华菊叶片局部横切面(×100);6硫华菊叶柄横切面(×40);Ep:表皮;Vb:维管束;Sp:海绵组织;Pp:栅栏组织;Ch:厚角组织;Ph:韧皮部;Xy:木质部
波斯菊叶片平均厚度为251.18μm,栅栏组织由1~2层长柱状细胞构成,厚度为98.92μm,细胞长轴与表皮垂直,排列疏松,含大量叶绿体;海绵组织厚度为105.34μm,细胞不规则,具胞间隙,含较多叶绿体。主脉由表皮、厚角组织、薄壁组织和维管束组成。厚角组织细胞达3层以上,主脉有3束外韧型维管束,呈三角形排列,中间一束较大(图4-1、2,表1)。硫华菊叶片厚度较波斯菊薄,平均厚度为230.49μm,栅栏组织由1层细胞构成,厚度为103.60μm,排列较紧密整齐,含大量叶绿体。海绵组织厚度为83.32μm,具发达胞间隙。主脉维管束数量及排列与波斯菊一致(图4-4、5,表1)。

表1 两种秋英属植物叶片显微结构比较
2.3.3 叶柄结构 波斯菊和硫华菊叶柄结构相似,横切面均近半圆形,近轴面平整,两侧凸起。叶柄由表皮、厚角组织、维管束和薄壁组织构成。表皮为一层排列紧密的圆形细胞组成,不含叶绿体。表皮下为厚角组织,波斯菊的厚角组织由2层细胞构成,硫华菊厚角组织细胞层数为3~4层。二者部分厚角组织细胞含叶绿体,使得叶柄横切面厚角组织部位呈现间断分布的绿色线条。薄壁组织细胞均较大,排列较疏松。维管组织均由5个椭圆形的维管束组成。中央的一个维管束最大,其它4个较小的维管束左右对称,靠近轴面的维管束最小。每一维管束包括木质部和韧皮部,木质部近轴面,韧皮部远轴面(图4-3、6)。
3 讨论
3.1 根的结构特点及生态适应性
波斯菊和硫华菊生长时期老根均具有次生结构。次生保护组织周皮的出现能更好地保护根内组织在根伸进土壤生长时免受土壤砂石的破坏,提高根穿透土壤的能力,同时还可以控制水分散失和防止病虫害、高温等不利因素对植物内部组织的损伤[7]。
此外,两种植物均具有发达的次生木质部。次生木质部中具有大量孔径不同的导管,有利于根系运输水分和无机盐,增强了植物适应干旱土壤环境的能力;次生木质部中纤维发达,发达的机械组织使根具有很好的强度和韧性,利于保护导管,保证水分运输的有效性和安全性。波斯菊与硫华菊的根虽形成次生结构,但根内部均具有发达的髓部。其中波斯菊的髓部占根直径的1/8,硫华菊的髓部占根直径的1/4。髓部由体积较大的薄壁细胞构成,成为营养物质和水分贮存的场所,为侧根及根毛的生长提供充足养分奠定了基础,提高根部抵抗逆境的能力。
3.2 茎的结构特点及生态适应性
波斯菊与硫华菊的茎均具有表皮毛,植物表皮毛一方面具有隔热的作用,使叶肉组织不会过热[8],另一方面可降低植物蒸腾作用,减少植物水分蒸发,是植物的抗旱性指标之一。通过观察,可发现二者表皮毛大小及结构有较大差异,波斯菊茎表面密布2~3个细胞构成的细小表皮毛,肉眼难辩;硫华菊表皮毛较粗大,呈弯钩状,多达20个细胞构成,长度可达1260μm。这两种表皮毛的具体生态作用还有待进一步研究。波斯菊与硫华菊茎均具有丰富的厚角组织及发达的髓部,体现二者茎具有较强的机械支持能力和抵御外界不利环境变化的能力。二者茎虽不粗,但茎内维管束均较丰富,其中波斯菊的维管束多达20个以上,在行使机械支持和输导作用方面具有良好的结构基础。
3.3 叶的结构特点及其生态适应性
叶是植物对逆境胁迫最敏感的部位,叶的形态结构是对环境适应性最直观的反映[9]。波斯菊与硫华菊均为异面叶,其叶片均具有角质层和表皮毛,栅栏组织发达,叶绿体丰富,气孔多分布于下表皮,这些结构基础共同构成阳生植物的典型特征[10]。其中,硫华菊的栅栏组织更为发达,其栅栏组织与叶厚度及海绵组织厚度的比值均较波斯菊大,发达的栅栏组织既可避免强光对叶肉的灼伤,同时说明其对光能的利用效率更高,其耐旱性也更强[11]。
猜你喜欢
小读者(2022年18期)2022-11-19
河北农业大学学报(2021年5期)2021-11-10
植物研究(2021年2期)2021-02-26
新疆农业科学(2020年9期)2020-10-13
诗选刊(2020年4期)2020-04-09
河南农业科学(2019年9期)2019-09-24
文学港(2018年1期)2018-01-25
天津农业科学(2015年11期)2015-12-03
儿童故事画报·发现号趣味百科(2015年7期)2015-10-23
林业与生态(2015年7期)2015-09-24
