
玉米秸秆施用对棕壤有机碳激发效应及温度敏感性的影响
2019-12-23 05:10李艾蒙裴久渤谢柠桧刘雨薇汪景宽
农业环境科学学报 2019年12期
李艾蒙,李 慧,裴久渤*,谢柠桧,刘雨薇,汪景宽
(1.沈阳农业大学土地与环境学院,土肥资源高效利用国家工程实验室,农业部东北耕地保育重点实验室,沈阳110866;2.沈阳农业大学林学院,沈阳110866)
农田土壤有机碳是土壤肥力的基础,其分解作为重要的土壤C释放和周转的驱动来源,对土壤肥力的固持和温室效应的形成具有直接作用[1]。玉米秸秆作为农田土壤重要的外源有机物,其归还影响土壤有机碳的合成与分解,进而影响着土壤培肥效果与温室效应的变化。在温室效应持续上升引发气候变暖的背景下[2],探讨秸秆添加如何影响土壤有机碳的激发效应及其对温度的敏感性对于揭示因气候变暖对土壤肥力产生的负效应具有重要意义[3]。
“激发效应”是Bingeman在1953年研究外源有机物质对土壤矿化的影响时提出的[4]。随后发现正负和大小差异,与外源有机物的数量和组成有关[5]。外源有机物进入土壤后,为微生物提供碳源,促进微生物数量上升,为满足自身的生存和发展,微生物在土壤中寻找营养元素,它们产生更多的胞外酶分解外源有机物并且对土壤原有机碳进行分解,从而导致正激发效应,也有研究学者称为“共同代谢”[6]。而负激发效应则被认为外源有机物输入之后,微生物会偏向利用外源有机物,对土壤原有机碳利用减少[7]。除了外源有机物输入影响激发效应外,温度和肥力水平的差异也是调控土壤有机碳激发效应最重要的因子,对土壤碳矿化的多少和快慢产生影响[8]。长期施肥处理影响了土壤肥力变化,与不施肥相比,施有机肥处理使土壤呼吸速率显著增加[9]。此外,土壤温度作为重要的环境因子,调控着土壤呼吸等一系列复杂的土壤生物化学过程[10]。通常情况下,增温能显著提高土壤呼吸速率[11],Meta 分析结果表明土壤增温0.3~6 ℃将导致土壤呼吸速率增加20%[12]。然而,外源有机物添加到不同肥力土壤后的有机碳矿化在不同温度下的激发效应,及其温度敏感性响应差异较少报道。
在复杂的农田土壤生态系统中,传统方法不能区分外源有机物对原土壤有机碳的激发效应及其对温度差异和肥力水平差异的响应,同位素方法的应用却可以解决此问题[13]。本研究利用13C 同位素示踪方法,在不同肥力土壤中添加13C标记的玉米秸秆,不同温度条件下进行室内培养,探究外源秸秆输入、温度差异、土壤肥力水平差异如何共同影响有机碳的损失,这对于揭示土壤有机碳周转过程对温室效应和农田管理措施的响应机理具有重要意义,为秸秆还田培肥地力效果的评估提供理论依据。
1 材料与方法
1.1 供试土壤
采自辽宁省沈阳农业大学棕壤长期定位试验站。试验地地势平坦,海拔75 m,属于北温带大陆性季风气候区,夏季高温多雨,冬季寒冷干燥。年平均气温7.5 ℃,年平均降水730 mm,≥10 ℃积温为3350 ℃,无霜期为153 d。土壤为发育在黄土性母质上的壤质棕壤(简育湿润淋溶土)。该试验站始建于1979 年,从1987 年开始布置肥料试验,每个处理3 次重复,每个重复小区面积为69.12 m2,一直坚持至今,均为玉米(当地常用品种)连作方式。试验开始前(1987 年)土壤(0~20 cm)有机质含量为15.6 g·kg-1,全氮为1.0 g·kg-1,全磷为0.52 g·kg-1,碱解氮为67.4 mg·kg-1,有效磷为8.4 mg·kg-1,速效钾为98.3 mg·kg-1,pH(H2O)为6.39。每年4 月25 日左右播种、施肥,并按常规进行田间管理;9月25日前后进行小区测产收割。选择其中2 种长期施肥处理土壤作为本研究中的高肥土壤、低肥土壤(表1),其中高肥土壤为每年施用含氮270 kg·hm-2的腐熟猪厩肥处理土壤,施用的腐熟猪厩肥有机质含量为150 g·kg-1左右,全氮为10 g·kg-1左右。所用肥料均作为基肥施入土壤[14]。低肥力土壤为不施肥处理土壤。
供试13C 标记秸秆于2016 年在田间利用原位脉冲标记获得。方法是在玉米生长最旺盛期进行13CO2原位脉冲多次(4~6 次)标记[15-16],玉米成熟后收获植株,105 ℃杀青30 min 后60 ℃烘干8 h。将烘干植物粉碎过0.425 mm 筛保存在密闭干燥的容器中备用。取少量粉碎后的植物样品再用球磨仪(Retsch MM200,德国)进行粉碎,利用元素分析-同位素比例质谱联用仪(EA-IRMS,德国)测定玉米秸秆含碳量和δ13C值。其含碳量为416.26 g·kg-1,全氮为11.60 g·kg-1,δ13C值为16.172%。
1.2 研究方法
试验处理:本试验共设计8 个处理:(1)25 ℃(微生物适宜温度)+高肥土壤+13C标记秸秆;(2)25 ℃+高肥土壤;(3)25 ℃+低肥土壤+13C 标记秸秆;(4)25 ℃+低肥土壤;(5)18 ℃(当地作物生长季平均温度)+高肥土壤+13C 标记秸秆;(6)18 ℃+高肥土壤;(7)18 ℃+低肥土壤+13C标记秸秆;(8)18 ℃+低肥土壤。
培养土壤预处理:2017 年9 月玉米收获后,在试验站所选的高肥和低肥处理土壤表层用土钻采集0~20 cm土层的土壤作为供试土样。每个重复小区按五点法随机采集后按四分法进行混合,作为1 个重复。将土样带回实验室挑除土壤中小石块和动植物残体,自然风干,过2 mm 筛备用。取相当于120 g烘干土质量的风干土于1 L 培养瓶中,调节含水量为田间持水量的40%,分别在25±1 ℃(微生物适宜温度)和18±1 ℃的恒温培养箱中预培养1周以恢复土壤微生物活性。
正式培养阶段:按照烘干土质量的5%添加13C标记的玉米秸秆,根据称重法定期补水调节含水量至田间持水量60%。每个处理3 次重复,随机排列。将装有土壤样品的培养瓶(用连接两根胶管的瓶盖封口,每根胶管外端均连接一个三通阀,一个用于连接换气装置、一个用于采集气体样品)分别置于25±1 ℃和18±1 ℃培养箱中进行避光密闭培养,每隔3~5 d 进行补水换气,为微生物提供充足的氧气。补水换气是在每次气体采集完毕后进行,步骤是先称质量补水再换气。补水后,立即密闭培养瓶,连接气体交换装置(一根胶管连接装置,另一根胶管打开三通阀用于瓶内气体交换),即连接一个气体换气泵,一端进入空气,中间连接碱柱吸收装置,可以吸收进入空气中的CO2,另一端连接的是培养瓶,以保证进入培养瓶的气体是没有CO2的空气,每次换气1 h 保证换气完毕后培养瓶的CO2浓度达到10 μL·L-1以下,关闭两根胶管三通阀,停止换气,继续培养试验。此外每个温度每个肥力处理另配备3 个环境CO2及其δ13C 误差校正瓶,即装120 g石英砂,按相应处理补水量进行补水。
样品采集与测定:在第5、60、140、300、360、450 d进行气体样品采集,将其中一根胶管的三通阀(另一根胶管三通阀关闭)连接螺旋注射器和气袋,进行气体样品采集。将采集后的气体样品用气相色谱-同位素比例质谱联用仪(GC-IRMS,德国)测定CO2含量及其δ13C值。
1.3 数据计算
土壤呼吸速率的计算:

式中:Rr为土壤呼吸CO2-C速率,μg C·g-1土·d-1;C为采集气体样品的浓度,μL·L-1;V为碱液吸收瓶中碱液上部空间体积,0.75 L;M为碳原子的摩尔质量,12 g·mol-1;W为培养瓶中烘干土质量,120 g;t为密闭时间,d。
秸秆碳对土壤呼吸的贡献比(Fmaize)的计算:

式中:δ13Csample和δ13Ccontrol分别代表秸秆添加和不添加处理CO2的δ13C 值(%),δ13Cmaize0代表培养前添加秸秆的δ13C值(%)。由此,秸秆添加处理中来自原土壤有机碳的CO2贡献比可计算为Fsoil=(100-Fmaize)×100%。相应地,来自秸秆碳和原土壤有机碳的CO2-C呼吸速率通过Rr与相应贡献比乘积得到。

表1 供试土壤的基本性质(2017年)Table 1 Basic properties of soil tested(in 2017)
激发效应(PE)及激发程度(RelativePE)的计算[17]:
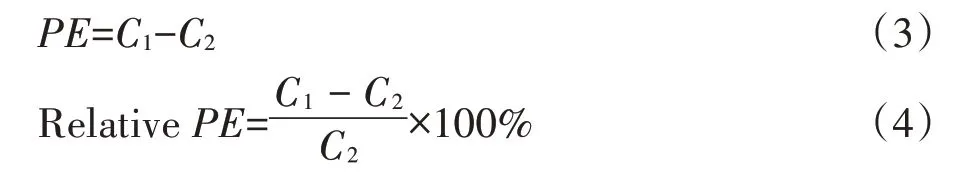
式中:C1和C2分别表示秸秆添加和不添加处理来自土壤原有机碳的呼吸速率。
土壤呼吸敏感性(Q)的计算:

式中:R0是T0参考温度(18 ℃)下呼吸速率,RT是T(25 ℃)温度下呼吸速率,Q是随温度升高的呼吸速率变化的敏感程度[18]。
此外,各指标累积量的计算通过按时间进行梯形积分而得到[17]。
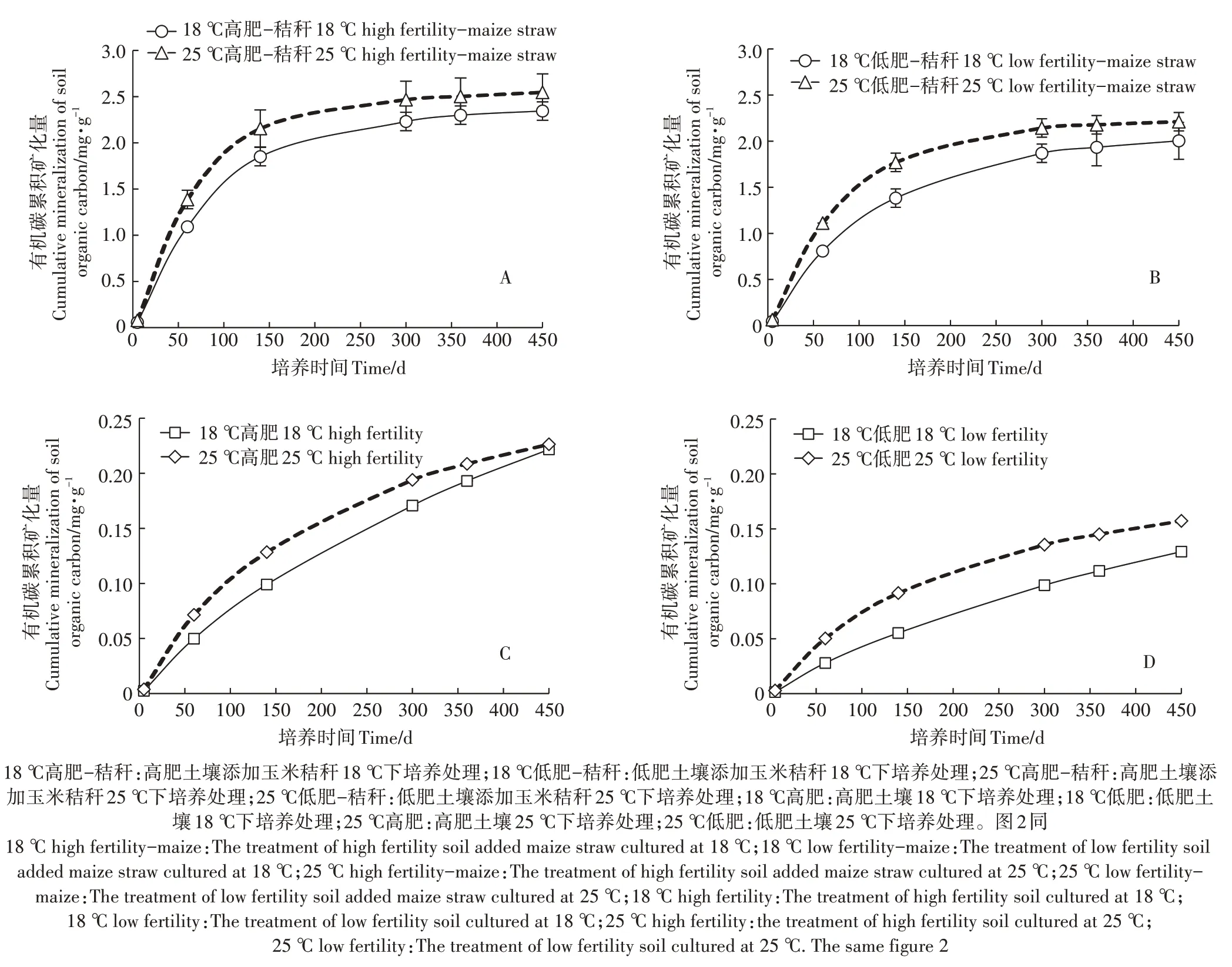
图1 棕壤有机碳累积矿化量Figure 1 Cumulative mineralization amount of organic carbon in Brown Earth
1.4 数据分析
采用Excel 2016 进行数据处理、绘制图表,SPSS 19.0 统计分析软件进行多因素方差分析和显著性检验(P<0.05),Origin 2018进行梯形积分计算。
2 结果分析
2.1 不同肥力棕壤的累积矿化量
由图1可见,25 ℃培养下有机碳的累积矿化量高于18 ℃的,并且高肥土壤的累积矿化量高于低肥土壤。秸秆添加显著促进了高肥土壤和低肥土壤的累积矿化,并且高肥土壤有机碳的累积矿化要高于低肥土壤。方差分析显示(表2),肥力水平、秸秆添加和培养温度对棕壤有机碳累积矿化量的影响均达到差异极显著水平(P<0.001),各因素之间也存在差异显著的交互作用(P<0.05)。
2.2 不同肥力棕壤的激发效应程度
秸秆添加后低肥棕壤激发效应程度变化大于高肥棕壤,高肥棕壤呈现0~300 d总体下降、后升高的趋势,在200 d以后呈现负激发效应先降低后升高,360 d后趋于平缓的趋势;而低肥棕壤0~300 d 也呈总体下降、后升高的趋势,但在140~350 d期间内呈现较长的负激发效应,之后表现出较平缓的正激发效应。图2C和图2D反映各采样时期的累积激发效应程度。培养期内,各个处理总体呈现出0~140 d 累积激发效应增加、随后降低(而低肥棕壤在360 d 后呈现增加)的趋势。方差分析表明(表2),培养温度和肥力水平显著影响了激发效应程度和累积激发效应(P<0.05),各影响因素之间存在着显著的交互影响作用(P<0.05)。
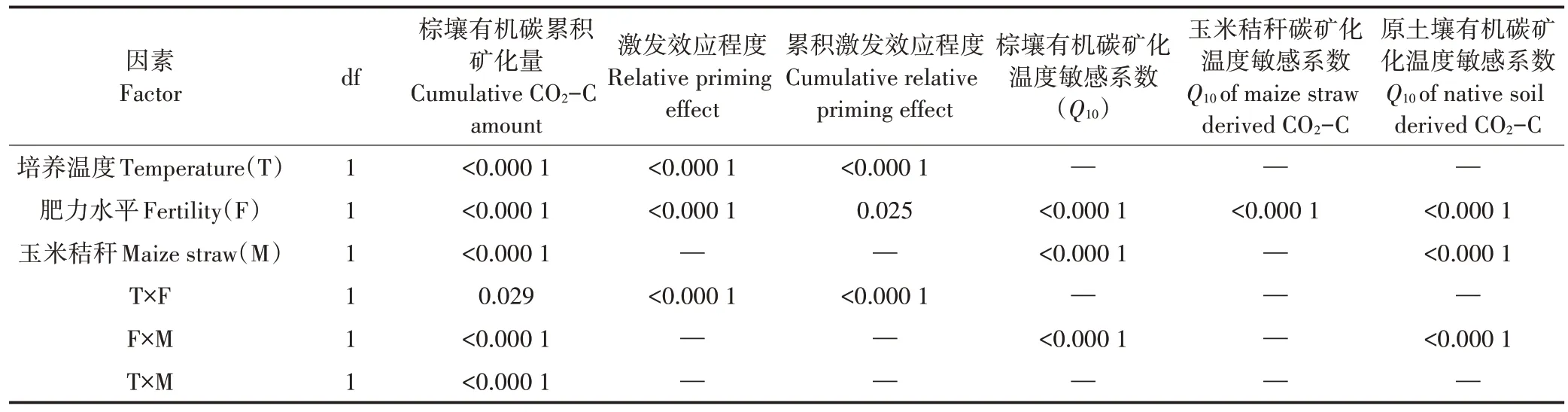
表2 多因素方差分析表(P值)Table 2 Multi-factor variance analysis(P value)
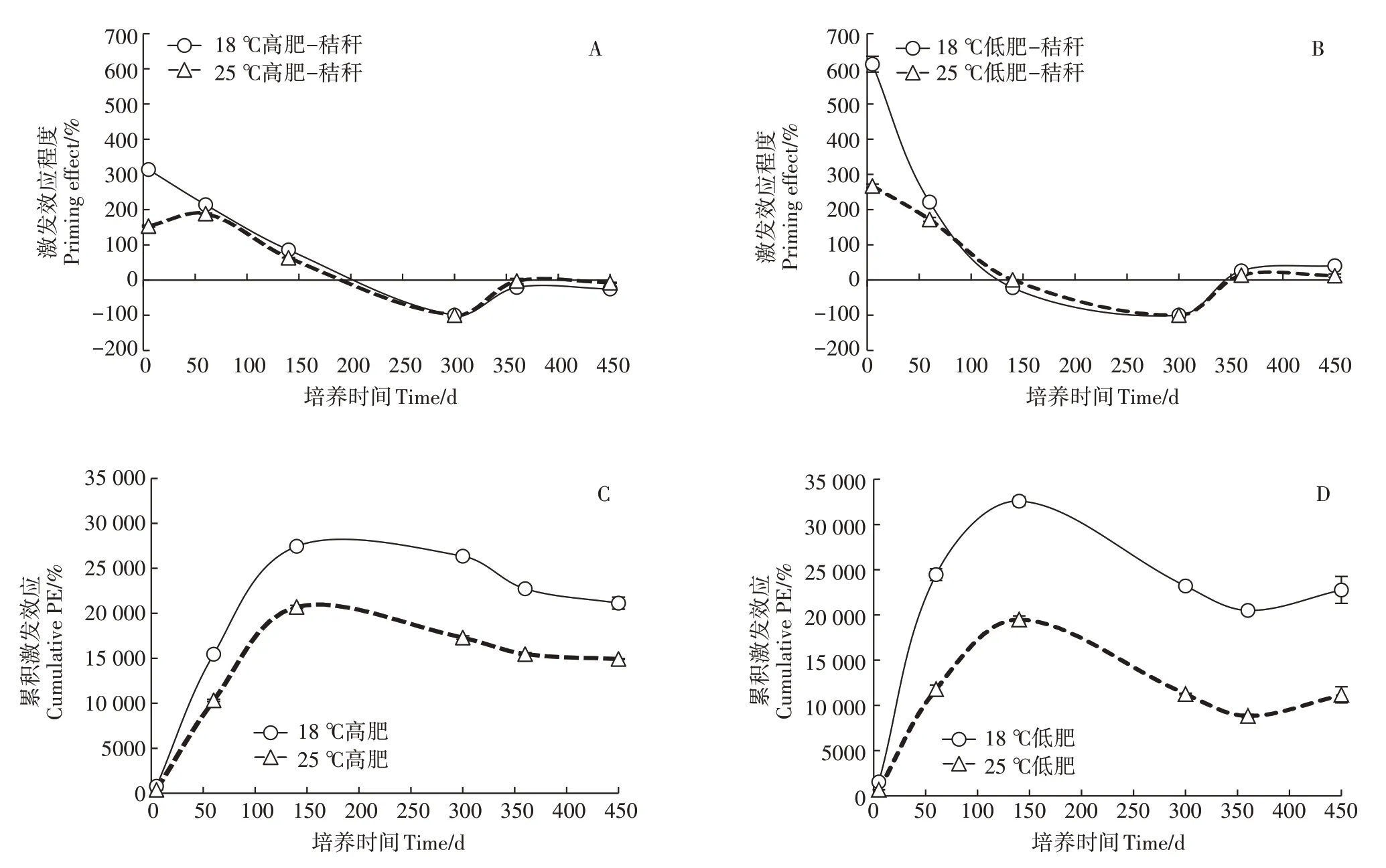
图2 秸秆添加后棕壤有机碳矿化的激发效应程度Figure 2 Relative priming effect of organic carbon mineralization in Brown Earth after maize straw addition
2.3 不同肥力棕壤呼吸的温度敏感性
培养期内,添加和未添加秸秆条件下高肥和低肥棕壤均呈下降趋势。未添加秸秆条件下,低肥棕壤的Q10高于高肥棕壤,且300 d 后均呈平缓趋势;而添加秸秆后两个肥力土壤的Q10较不添加秸秆均表现为降低趋势,呈现出0~140 d 高肥低于低肥棕壤,而后低肥低于高肥棕壤的变化趋势”,并在300~360 d 呈现先升高再降低的趋势(图3)。方差分析显示(表2),肥力水平和玉米秸秆添加对棕壤Q10的影响均达到差异极显著水平(P<0.001),各因素之间也存在差异极显著的交互作用(P<0.001)。
2.4 不同肥力棕壤有机碳矿化中秸秆来源碳和原土壤有机碳的温度敏感性
培养期内高肥和低肥棕壤中玉米秸秆来源碳矿化Q10呈总体下降趋势(图4A),其中140 d 前呈现出低肥棕壤高于高肥棕壤的趋势,而后表现为相反趋势,且300 d后呈现出略升高后降低的变化。图4B表示添加秸秆后高肥和低肥棕壤原土壤有机碳矿化Q10变化,总体呈现先增加后降低趋势,其中300 d以前呈现低肥棕壤显著高于高肥棕壤的趋势,但最高值的出现晚于高肥棕壤,而300 d 后呈现略升高后降低的趋势,且低肥棕壤低于高肥棕壤。方差分析显示(表2),肥力水平对玉米秸秆碳矿化Q10的影响达到差异极显著水平(P<0.001),并且秸秆添加后对原土壤有机碳矿化Q10的影响及交互作用也达到差异极显著水平(P<0.001)。
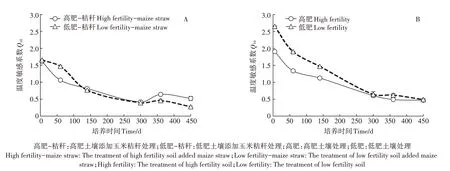
图3 棕壤有机碳矿化的温度敏感系数(Q10)Figure 3 Q10 of soil respiration
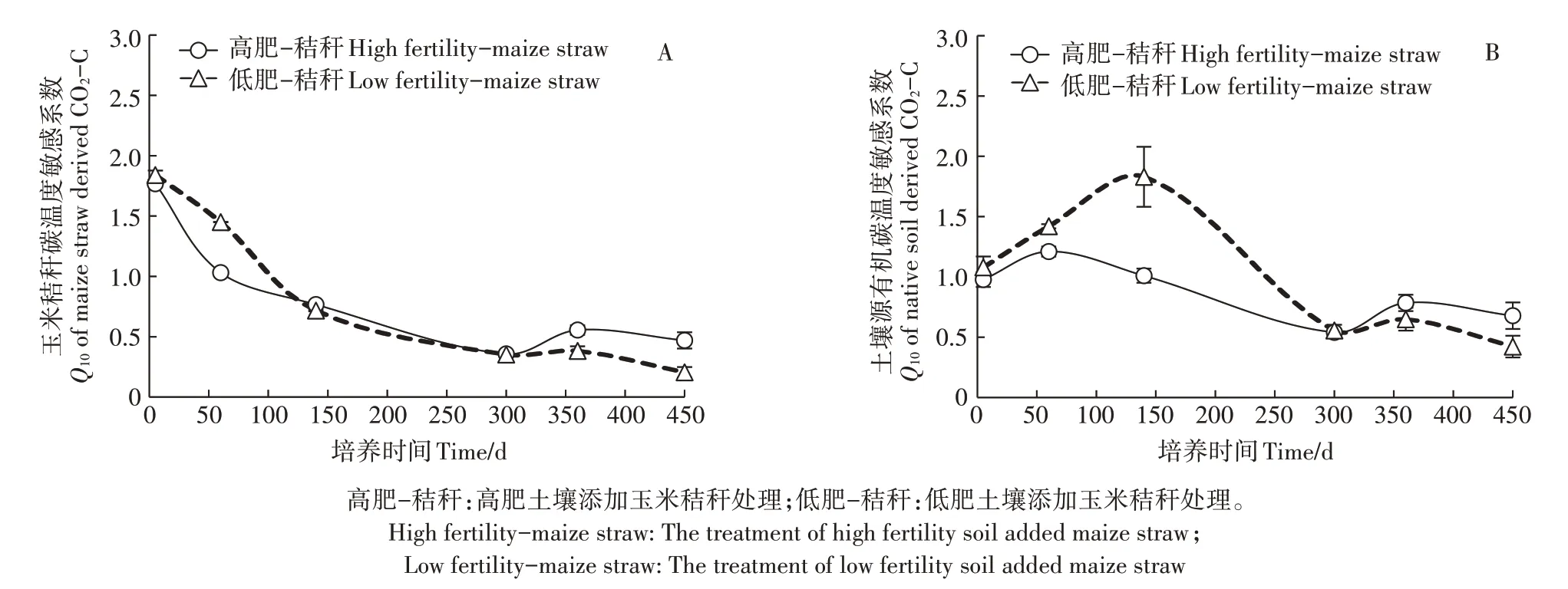
图4 玉米秸秆碳和原土壤有机碳矿化的温度敏感系数(Q10)Figure 4 Q10 of maize derived-and native soil derive CO2-C
3 讨论
3.1 秸秆添加后肥力和温度对棕壤有机碳激发效应的影响
长期定位研究发现,土壤碳输入量增大并不一定导致土壤有机碳储量的增长,因为土壤碳排放量也随之提高[19]。这种外源有机质输入在短时期内改变土壤有机碳矿化过程的现象称为激发效应[20]。肥力水平的差异(土壤有机碳水平差异)将引起微生物对新添加秸秆碳和土壤有机碳分解的竞争,从而影响土壤有机碳的激发效应,即低肥土壤有机碳含量低、养分相对缺乏,使微生物长期处于“饥饿”状态[21],微生物活动所需的养分受到限制[22],而外源玉米秸秆的添加使土壤中这部分微生物活化,刺激微生物分泌更多的胞外酶,同时加速对土壤原有机碳的矿化,产生正激发效应。随有机物质分解速率减慢,激发效应的程度也降低[23-24]。因此,本研究中低肥土壤中添加玉米秸秆后,前期产生的正激发效应程度呈现逐渐降低趋势。而随着有机物料的分解,土壤有机碳激发效应的变化呈现“U”型变化趋势,即在初期与后期呈现正激发效应,但在中期呈现负激发效应[25]。这与本研究培养各时期表现的结果基本一致,而低肥土壤负激发效应的持续时间长可能是由于基础有机碳含量明显偏低,秸秆碳进入土壤后其有机物质分解、释放,相对增加了土壤有机碳含量,减缓了其损失[26]。温度差异是影响土壤有机碳矿化的另一个重要因素,影响了微生物活性、土壤有机碳矿化速度,进而影响土壤有机碳激发效应的变化[27]。这种变化将通过激活微生物活性来释放出更多胞外酶来促进原有SOC 分解[28];而通过升高温度降低SOC 的活化能也会使SOC 分解加快[29]。这两种效应处于此消彼长的状态,加强其中一种作用机理,必然会削弱另一种。因此土壤有机碳激发效应会出现18 ℃培养条件下高肥土壤中添加玉米秸秆后0~300 d 激发效应程度比25 ℃高,而300~450 d相反,18 ℃培养条件下低肥土壤添加玉米秸秆后0~140 d 和360~450 d 激发效应的程度比25 ℃高,而140~360 d相反。内在的微生物变化调控激发效应的机理需要今后深入研究。
3.2 秸秆添加后肥力和温度对棕壤有机碳矿化温度敏感性的影响
温度敏感性(Q10)能够反映土壤有机碳矿化受温度变化的影响大小,且因影响因素的改变会引起响应结果的不同[30-32]。当培养温度升高后,土壤有机碳分解速率增快,表现为对增温的响应更敏感,Q10更高[33]。此外,底物的差异也会引起Q10对温度变化的响应。Verburg 等[34]在控制温度和水分条件的培养实验中,发现土壤呼吸存在季节变异,原因是底物供应的季节变异。表明底物供应对温度敏感性有着重要影响,当土壤中底物供应减少时,土壤微生物会降低活性甚至死亡,Q10会随之降低。本研究结果也显示添加玉米秸秆后在各个时期均引起土壤有机碳矿化的温度敏感性的变化(图3A 和图4A),而140 d和300 d 是受高、低肥影响下土壤有机碳矿化温度敏感性发生变化的重要时期。将反映出易分解和难分解有机物质含量的变化引起的土壤有机碳矿化的温度敏感性的变化[23]。Gren 等[35]从微生物酶促动力学方向研究了温度敏感性与底物质量的关系,认为复杂的有机化合物一般具有较低的分解速度和较高的活化能,随着土壤有机质分子量和分子结构复杂性的增加,促使其发生生化反应所需的能量也增加,因而对温度的敏感性也相应地增加。而培养300~450 d 微生物可能转向复杂的有机化合物,高肥土壤Q10大于低肥土壤,是因为相对于低肥土壤,添加玉米秸秆的高肥土壤复杂的有机化合物较多,土壤呼吸速率增强,Q10值升高,土壤有机碳的难分解组分比易分解组分具有更高的温度敏感性,这与Knorr等[36]结论一致。
3.3 肥力和温度对玉米秸秆来源碳和原土壤有机碳矿化的温度敏感性的影响
土壤有机碳矿化和激发效应的变化差异,主要由玉米秸秆碳和原土壤有机碳的消减变化引起,肥力和温度是影响其敏感性变化的主要因素。图4A 中0~140 d 低肥棕壤玉米秸秆碳矿化Q10高于高肥,是由于长期不施肥土壤中养分含量较低,加入新碳后,温度的升高激发了微生物的利用,Q10增大,表明高肥土壤140 d 前有利于稳定秸秆碳的温度敏感变化。而300 d 后高肥和低肥土壤呈现略升高后降低的趋势,是由于微生物转向土壤中较难分解状态的有机质,而高肥土壤中较多的难分解物质会引起土壤呼吸速率增强,Q10升高,随底物的消耗,土壤呼吸速率降低,Q10又呈现下降趋势[36]。对于原土壤有机碳矿化的温度敏感性变化主要是温度影响微生物利用秸秆碳和原土壤有机碳之间的差异所引起。受肥力水平影响,0~300 d 培养期内低肥土壤微生物更易于被温度和外源有机碳影响,从而影响原土壤有机碳的分解而引起Q10强烈变化,随着外源有机碳被微生物逐渐消耗,原土壤有机碳Q10的变化逐渐与外源有机碳Q10的变化趋于相似,这也与Michaelis-Menten 方程的原理解释基本一致[37]。
4 结论
本研究结果总体反映出室内培养条件下,棕壤有机碳的矿化显著受秸秆添加、肥力水平(有机碳的差异)、温度高低的综合影响。
(1)肥力、温度的升高均会增加棕壤有机碳的累积矿化量。
(2)18 ℃培养条件下棕壤累积激发效应程度高于25 ℃,其中低肥棕壤的总体变化大于高肥棕壤。
(3)玉米秸秆碳加入,降低了高肥和低肥棕壤有机碳矿化的温度敏感性,表现出与秸秆碳温度敏感性变化较为一致的变化趋势,而原土壤有机碳矿化的温度敏感性呈现较大差异变化,140 d和300 d是其变化的重要转折时期。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
口腔医学(2021年10期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
能源工程(2021年3期)2021-08-05
浙江农林大学学报(2019年6期)2019-11-13
医学研究杂志(2015年9期)2015-07-01
农业科技与装备(2014年7期)2014-11-28
