
大洲岛海杧果群落及其物种多样性特征
2019-12-19 02:13吕林玲陈慧李伟杰杨海建戴波黄良明郑希龙
热带作物学报 2019年11期
吕林玲 陈慧 李伟杰 杨海建 戴波 黄良明 郑希龙

摘 要 通过野外实地调查对海南省万宁市大洲岛的海杧果群落特征及其物种多样性进行研究,以期为海杧果的种质保护、海防林物种的合理搭配及红树林生态恢复提供参考。结果表明:调查样方内有维管束植物62种,隶属于37科58属。乔木层的优势种为海杧果,重要值为36.45%;灌木层的优势种为矮紫金牛,重要值为8.31%;草本层的优势种为观音草,重要值为25.53%。种子植物区系成分以热带成分占优势,泛热带分布的科属最多,分别为27科、17属。群落物种丰富度Margalef指数、Shannon-Wiener指数、均匀度Pielou指数排序均为灌木层>乔木层>草本层,优势度Simpson指数为灌木层>草本层>乔木层。本调查明确了大洲岛海杧果群落的伴生物种及该群落特征,可为红树林生态恢复及海防林建设提供参考。
关键词 大洲岛;海杧果;红树林;区系特征;物种多样性
中图分类号 Q948.15 文献标识码 A
Abstract In this study, the features and species diversity of the Cerbera manghas communities in Dazhou Island, Wanning, Hainan were studied based on field investigation, in order to provide references for the germplasm conservation, the reasonable matching of coast shelter-forest species and the ecological restoration of mangrove forest. There were 62 species in 58 genera and 37 families of vascular plants in the quadrats. Cerbera manghas, Ardisia humilis, and Peristrophe bivalvis occupied the predominant position in the tree layer (36.45% of Importance Value (IV)), shrub layer (8.31% of IV) and herb layer (25.53% of IV) respectively. Flora of seed plant was dominant with tropic zone. The pantropic type was the most widely distributed form, with 27 families and 17 genera. The Margalefs index, Shannon-Wiener index and Pielous index order of all layers in the community was shrub layer>tree layer>herb layer. The pattern of the Simpson index was shrub layer>herb layer>tree layer.
Keywords Dazhou Island; Cerbera manghas; mangrove forest; flora characteristics; species diversity
DOI 10.3969/j.issn.1000-2561.2019.11.024
海杧果(Cerbera manghas L.)为夹竹桃科Apocynaceae海杧果属Cerbera L.植物,在海南又被称为黄金茄、牛金茄、黄金调、牛心荔。其因果实似芒果,生于海岸边,故得名海杧果,分布于中国广东省、海南省、广西壮族自治区、台湾省;柬埔寨、印度尼西亚等东南亚国家,日本、澳大利亚也有分布[1-3]。海杧果的树皮、叶、乳汁多有药用,制成药剂可用于催吐、下泻、堕胎,有一定的药用价值,但服用量过大能致死,且其果皮、种子有剧毒,应慎用[4-5];此外,国内外已有许多学者对海杧果的化学成分及生物学活性进行了研究,发现其具有一定的抗癌作用[2, 6-10]。红树林是海岸带重要的植被类型之一,有防风护堤、维护海岸生态平衡等多方面作用[11],而海杧果是半红树植物,其耐盐碱、喜热,因此多生于高潮可及的海岸带,亦有防风护林作用。陆彦盼等[12]为保护海杧果种质资源,对琼北地区的海杧果群落进行了调查,并对种群内各种表型性状进行分析,但小面积的离岸岛与面积广阔的海南本岛相比,环境差异较大,目前对海南省本岛外离岸岛屿的海杧果群落的研究还较少。群落多样性是生物群落的重要特征,反映群落自身特征及其与环境之间的相互关系[13]。研究植物群落的多样性是研究植物群落在组成结构、功能和动态方面表现的丰富多彩的差异[14],而物种丰富度、多样性指数和均匀度指数是表征群落特征的重要指标,在反映植物群落的结构类型、演替阶段和稳定程度等方面具有重要意义[15]。因此本文通过分析处于海南省万宁市东南部的大洲岛上的海杧果群落多样性特征,了解其种群的生长规律,以期为海杧果的种质保护、海防林物种的合理搭配及红樹林生态恢复提供基础的参考依据。
1 研究区域环境概况
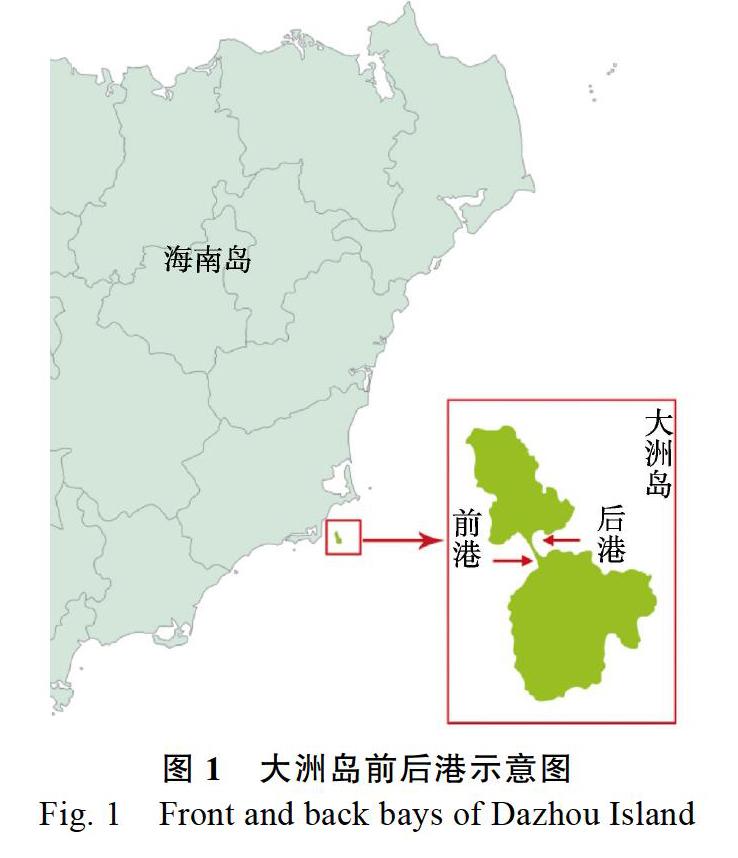
大洲岛又名燕窝岛,也称独洲岭、独珠岭,地处海南省万宁市东南部,位于东经1100126~ 1102940,北纬183919~184126,距万宁县城15 km,与乌场港隔海相望(图1)。大洲岛为海南省沿海面积最大的离岸岛屿,海岛陆地面积4.2 km2,主峰高约289 m。该区域属热带海洋性季风气候,年平均气温24.8 ℃,雨量充沛,年平均降水量2166 mm,阳光充足,年平均日照时数达2108.9 h。大洲岛植被丰富,且因其较早设立保护区,岛内植被保存较为完好,植被类型多样,以常绿季雨林为主,有少部分沙生及沼生植被,植物区系则以热带成分居多。
中国的红树林主要分布于海南、广东、香港、澳门、广西、福建、台湾及浙江等地[11],海杧果仅在中国南方少数沿海省份有栽培。陆彦盼等[12]调查发现,海南省其他市县的海杧果群落多生长于农田田埂、池塘边上及滨海天然林地,且认为干扰情况较为严重,这与大洲岛海杧果的生长环境有一定差异。大洲岛沿岸地带多为耐风蚀及海水侵蚀的花岗岩,常形成崎岖的石滩,因此岩石的裸露程度较高,多在70%以上,而大洲岛的海杧果群落沿该岛的海岸带分布,生长在海拔较低处。本文主要调查在大洲岛前、后港地区集中分布的海杧果群落,分布区的最高海拔为61 m,最低海拔约3 m,大部分位于高潮可及地带。
2 方法
2.1 调查方法
野外调查工作于2017年2—4月进行。在大洲岛进行全面踏查后,选择海杧果典型分布的地域,根据海岛实际地理情况及海杧果生长状况,设置样方进行调查。其中,设置了5个10 m×10 m的乔木样方。同时,在每个乔木样方的中心设置1个5 m×5 m的灌木样方,在乔木样方的4个角落各设置1个1 m×1 m的草本样方。样方概况见表1。
根据方精云等[13]的植物群落清查的主要内容、方法和技术规范,用“每木记账法”记录:(1)胸高直径(Diameter at Breast Height, DBH)≥3 cm的乔木树种的种名、胸径、株高以及冠幅;(2)灌木树种的株数、株高以及盖度;(3)草本植物的种数、盖度和多度。
2.2 数据处理
2.2.1 群落重要值 群落重要值(Important Value, IV)是衡量不同群落间某一物种重要性的一个定量指标,本文物种的重要值参考方精云等[13]的计算方法进行统计,其中多度为某一物种在样方中统计得到的株数;频度是指某一物种在所有样方中出现的频率;优势度指某一物种的胸高断面积;盖度指某一物种地上部分的垂直投影面积占样方面积的百分比。
(1)乔木层重要值计算公式:
(2)灌木层及草本层重要值计算公式:
2.2.2 植物区系 根据李锡文[16]关于中国种子植物区系的划分和吴征镒[17]关于中国种子植物属的分布区类型划分,统计群落中植物科、属的分布区类型。
2.2.3 群落多样性指数 本文采用Margalef指数[18]、Simpson指数[19]、Shannon-Wiener指数[20]、Pielou指数[20]4个指标来分析海杧果群落的α多样性。
(1)丰富度Margalef指数计算公式:
(2)优势度Simpson指数计算公式:
(3)Shannon-Wiener多样性指数计算公式:
(4)Pielou均匀度指数计算公式:
式中,S为样方的植物种类总和;Pi为种i的个体数占所有种个体数的比率;N为样方所有物种的个体数之和。
3 结果与分析
3.1 群落特征
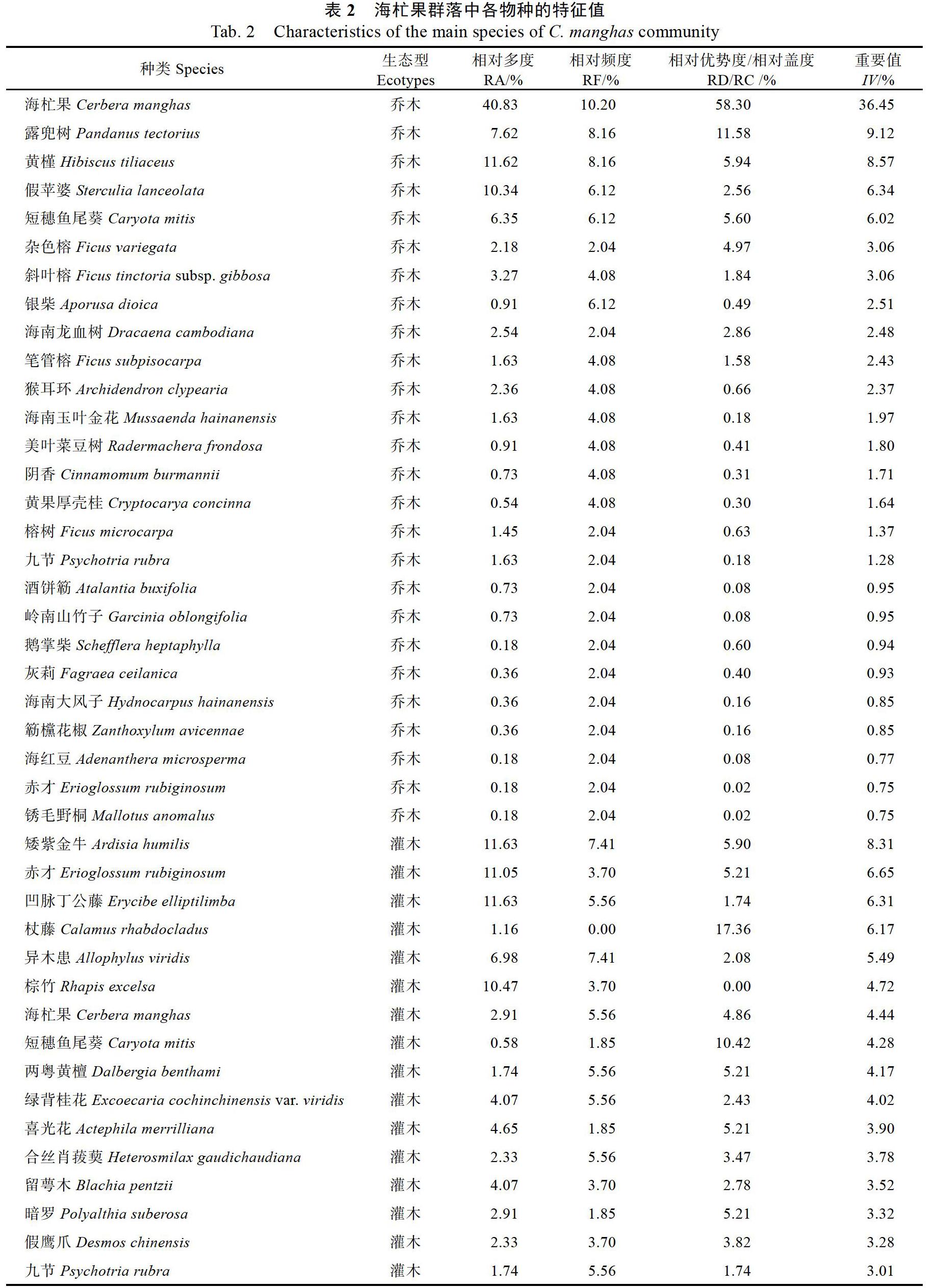
3.1.1 物种组成 调查发现,海杧果集中分布在大洲岛前、后港两岸的低海拔沿岸,群落中共有维管束植物62种(含种下分类,见表2),隶属于37科58属。其中,蕨类植物1科1属1种,单子叶植物8科12属12种,双子叶植物28科45属49种。
3.1.2 科的组成特点 5种以上(含5种)的有1个科,占总科数的2.70%,为大戟科Euphorbiaceae,共6种;桑科Moraceae、芸香科Rutaceae各有4种;棕榈科Palmae、爵床科Acanthaceae、禾本科Gramineae、番荔枝科Annonaceae、无患子科Sapindaceae各3种,共占总数的23.81%;含2种的科有4科;仅含1种的科有25科,占总科数67.57%,共占总种数的39.68%。
3.1.3 属的组成特点 仅含1种的属有56属,占总属数的96.55%,因此组成该群落的物种科属比较分散,含单种的科较多,含单种的属最多。含2种以上的属仅有榕属Ficus、紫金牛属Ardisia;其中榕属植物最多,有4种,分别为榕树Ficus microcarpa、杂色榕F. variegata、斜叶榕F. tinctoria subsp. gibbosa、笔管榕F. subpisocarpa。
3.1.4 群落特征值 乔木层有植物26种,551株;灌木33种,172株;草本13种。此外,记录到的层间植物有16种,如两面针Zanthoxylum nitidum、海南青牛胆Tinospora hainanensis、凹脉丁公藤Erycibe elliptilimba、赤苍藤Erythropalum scandens等。群落外貌完整,树冠连片生长,群落内植物层次分明。在乔木层中,海杧果占有主要地位,重要值为36.45%。其中,南岛海杧果植株较高,为3~5 m。而北岛海杧果群落由于处于风口处,植株较南岛的矮,为2~4 m高。露兜樹Pandanus tectoriu和黄槿Hibiscus tiliaceus为群落中地位仅次于海杧果的乔木,重要值分别为9.12%、8.57%。灌木层中无明显优势种,矮紫金牛Ardisia humilis、赤才Erioglossum rubiginosum分布较多,其中矮紫金牛的重要值最高,为8.31%。由于林下郁闭度高,草本种类较少,在群落中零散分布,主要有长叶肾蕨Nephrolepis biserrata、百足藤Pothos repens、草豆蔻Alpinia hainanensis、厚叶崖爬藤Tetrastigma pachyphyllum、马?儿Zehneria indica 5种。
3.2 区系特征
3.2.1 种子植物科的区系成分分析 海杧果群落中,共有种子植物61种,隶属于36科57属,其中科的分布区类型有6个(表3)。世界分布科有3科,分别为百合科Liliaceae、禾本科、旋花科Convolvulaceae,占該群落中所有种子植物科的8.33%;北温带分布科仅有紫草科Boraginaceae,占该群落中所有种子植物科的2.78%;其余32科为热带分布科,占总科数的88.89%;说明海杧果群落的植物区系类型以热带区系为主,其中又以泛热带分布的科最多,有27科,占该群落中所有种子植物科的75.00%。
3.2.2 种子植物属的区系成分分析 统计得到海杧果群落中种子植物属的分布区类型有8个(表3)。热带分布成分占优势,共55属,占该群落种子植物属的96.49%,其中泛热带分布属最多,有17属,占该群落种子植物属的29.82%;其次为热带亚洲至热带大洋洲分布的属,有14属,占24.56%;旧世界热带分布属也有10属,占17.54%,排在第3位。地中海区、西亚至中亚分布仅有1属,为木犀榄属Olea,中国喜马拉雅(SH)分布仅有1属,为矢竹属Pseudosasa。属级区系类型表明,海杧果群落中种子植物属的区系成分以热带成分占优势,也存在少量其他分布类型。
3.3 群落中不同层级植被的物种多样性分析
由图2可知,大洲岛海杧果群落中各层的物种丰富度Margalef指数(E)、Shannon-Wiener多样性指数(H')表现为:灌木层>乔木层>草本层,说明群落各层中灌木层的物种数最多,草本层的物种数最少,且灌木层优势地位较乔木层明显,而草本层优势地位最低,这与灌木层种类及数量多有关,而且灌木层存在一定数量的乔木种类的幼苗,表明此群落有实生苗不断补充更新,属于增长型群落。
由图3可知海杧果群落中各层的优势度(D)指数大小为:灌木层>乔木层>草本层,结果进一步说明灌木层优势地位较乔木层明显,且该层植物分布最均匀,最小的为草本层,这是因为群落荫蔽度高,林下草本少,多数样方内无草本存在;Pielou均匀度指数(Jsw)大小为:灌木层>草本层>乔木层,乔木层的均匀度指数最小,说明其在群落中分布不均,但是其林冠连接紧密。
4 讨论
4.1 群落组成特征分析
大洲岛的海杧果群落其群落较为完整,群落中层次分明、结构合理,乔木、灌木、草本及层间植物均占一定比例,人为干扰较少。其植株高度等高或远高于其附近的其他物种,因此树冠层能吸收到充足阳光,生长旺盛,但树冠连接紧密而使林下荫蔽度较高,造成林下草本较少。本次记录到海杧果群落内有维管束植物62种,分别属于37科58属。
据杨盛昌等[11]、何斌源等[21]统计,中国的原生红树植物分别为38种和36种(含1变种),但是并未详细记录红树植物群落的伴生植物。在海南省本岛的红树林里,海杧果一般为作为伴生物种小面积存在,且其所在的红树林物种一般以桐花树、木榄、卤蕨等真红树植物为主,非红树物种较少[22-24],但大洲岛上分布的海杧果面积较大,独立成林,且林下植物丰富。大洲岛海杧果群落为自然群落,属于半红树林植被型,其生长环境长年风力较大,每年7—11月偶有台风侵袭,但该群落仍生长茂盛,说明群落的抗风能力较强。除了乔木层的优势种海杧果外,其重要的伴生物种黄槿及露兜树为海南省海岸带常见植物,抗风能力亦极强。红树林的主要作用之一便是作为沿海地区的缓冲带,减少极端天气造成的损失,半红树植物。明确本群落的物种组成,可为进行红树林群落的修复或人工栽培时的合理布置各层级物种、搭配伴生物种提供一定的参考。
4.2 群落区系特征
大洲岛海杧果群落植物区系类型比较简单,科、属的分布类型皆以热带成分占优势,其中热带性质分布科占88.89%,热带性质分布科属占96.49%,其中又以泛热带分布的科属最多,分别有27科、17属,其余分布类型较少;群落中种类较多的大戟科、桑科、芸香科、棕榈科、爵床科及榕属、紫金牛属皆为泛热带分布类型;此外,大戟科、桑科在整个大洲岛亦是优势科,分布的种类皆有20种以上,对大洲岛植物区系的发展具有重要作用。其区系特征与蔡开朗等[25]在三亚市铁炉港红树林自然保护区的调查结果一致,亦是热带区系为主,其中又以泛热带成分占优,并且有少数温带分布型。此外,区系调查结果与张永夏等[26]、秦新生等[27]、张荣京等[28]在海南地区开展的其他非红树植物群落区系的调查结果亦相似,皆以热带性质分布型为主;总体看来,大洲岛海杧果群落区系与海南省本岛植物区系一致,这与大洲岛地处热带北缘地区的地理位置相符。群落中无中国特有属分布,反映了大洲岛植物区系分化的历史较短[29]。
4.3 群落多样性特征
一个群落的生态优势度越小,均匀度越大、丰富度指数越大,则该群落的多样性就越高[30]。本次调查发现,群落中除草本层外,乔木层、灌木层的Shannon-Wiener物种多样性指数均大于2.0,而群落中灌木层的各项指数均最高,这是因为灌木种类中还包含了乔木幼树,而且大洲岛常年大风,乔木层受台风等因素的影响,植株数量被制约,而林下草本由于群落荫蔽度高这一原因,种类及数量都较少,因此其各项指数较低,但草本层的Pielou均匀度指数较乔木层高,可能是因为草本样方中多数无草本分布,因此造成平均度高这一现象。宋贤利等[31]总结前人研究,认为群落的物种多样性指数、均匀度指数在一定意义上能够说明群落的结构类型。本研究中群落层次的多样性格局大致表现为灌木层>乔木层>草本层;海杧果在乔木层与灌木层均有分布,表明海杧果群落可正常更新演替,该群落正处于属于增长演替阶段,尚未发展成顶级群落。
4.4 大洲岛与海南琼北地区海杧果群落特征比较
据陆彦盼等[12]的调查,琼北地区的海杧果多生长在农田、池塘边及滨海林地等土层较厚的地方,而大洲岛上的海杧果多生长于岩石裸露程度达70%以上的区域,部分生于海岸沙地,且處于台风或大风时常肆虐的海岸带区域;这说明海杧果对环境的适应性较强,且有良好的抗风能力。从分布海拔来看,琼北地区及大洲岛地区的海杧果种群均主要生长在海拔较低的地区,但在大洲岛,海拔分布上限可达61 m。大洲岛上的海杧果植株长势良好,平均高度约为4.5 m,而琼北各地区的植株高度差异较大,主要与其立地条件相关,立地条件好的情况下树高甚至可达12 m。此外,海南省琼北地区的海杧果群落由于未得到重视,遭受到严重的人工干扰及破坏,因此其种群破碎化严重,植株分布不均匀,成片分布的种群较少;而在大洲岛上分布的海杧果群落虽然不多,但是其种群生长较为集中,这与其生长于离岸岛,人为干扰较少,且位于保护区内,受到一定的保护有关,上述现象说明人工保护对其种群发展、恢复极其重要。
海南省红树林比其他省区的红树林植物生态多样性更加丰富[32],真红树专一长于潮间带,而半红树既能生于潮间带,亦能生长于非盐渍土上[33],而海杧果是乔木类型的半红树植物,其优良的环境适应性、良好抗风的性能使其可以作为海防林替代树种,且其株形优美,观赏价值高,作为海岸园林景观树种沿海岸带种植亦是不错的选择;因此海杧果种群的保护工作应该得到重视,目前海南省本岛的海杧果群落受到的关注度低,种群的许多特性仍有待进一步研究。
致 谢 大洲岛国家级海洋生态自然保护区的钟昌芬、吴英弘、卓云川等人对我们在大洲岛上的生活及工作提供了帮助,福建农林大学黄柳菁博士为本文的写作提供了建设性的意见,特致谢意!
参考文献
[1] 广东省植物研究所. 海南植物志:第3卷[M]. 北京: 科学出版社, 1974: 228-229.
[2] 李海燕. 海杧果化学成分研究[D]. 海口: 海南大学, 2010.
[3] Flora of China Editorial Committee. Flora of China: Vol.16[M]. Beijing: Science Press and Missouri Botanical Garden Press, 1995,164-165.
[4] 中国科学院中国植物志编辑委员会. 中国植物志:第63卷[M]. 北京: 科学出版社, 1977: 33-35.
[5] 陈封怀. 广东植物志:第1卷[M]. 广州: 广东科技出版社, 1987: 431-432.
[6] Gaillard Y, Pepin G. Poisoning by plant material: review of human cases and analytical determination of main toxins by high performance liquid chromatography (tandem) mass spectrometry[J]. Journal of Chromatography B: Biomedical Sciences and Applications, 1999, 733(1-2): 181-229.
[7] Wang Y S, He L, Wang Q J. Study on the chemical composition and its pharmacology of the medicinal mangrove plant[J]. Chinese Journal of Marine Drugs, 2004, 23(2): 26-31.
[8] 张小坡, 裴月湖, 刘明生, 等. 海杧果叶中有机酸类成分研究[J]. 中草药, 2010, 41(11): 1763-1765.
[9] 李海燕, 王茂媛, 邓必玉, 等. 海杧果茎的挥发性成分研究[J]. 时珍国医国药, 2010, 21(7): 1676-1677.
[10] 王祝年, 李海燕, 王建荣, 等. 海杧果化学成分与药理活性研究进展[J]. 中草药, 2009, 40(12): 2011-2014.
[11] 杨盛昌, 陆文勋, 邹 祯, 等. 中国红树林湿地: 分布、种类组成及其保护[J]. 亚热带植物科学, 2017, 46(4): 301-310.
[12] 陆彦盼, 吕 冰, 赵 威, 等. 海南琼北地区海杧果种质资源调查与分析[J]. 林业资源管理, 2015, 6: 160-164.
[13] 方精云, 王襄平, 沈泽昊, 等. 植物群落清查的主要内容、方法和技术规范[J]. 生物多样性, 2009, 17(6): 533-548.
[14] 黄忠良, 孔国辉, 何道泉. 鼎湖山植物群落多样性的研究[J]. 生态学报, 2000, 20(2): 193-198.
[15] 王伯荪, 余世孝, 彭少麟, 等. 植物群落学实验手册[M]. 广州: 广东高等教育出版社, 1996.
[16] 李锡文. 中国种子植物区系统计分析[J].云南植物研究, 1996, 18(4): 363-384.
[17] 吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究, 1991(SIV): 111-139.
[18] Magurran A E. Ecological diversity and its measurement[M]. New Jersey: Princeton University Press, 1988.
[19] Simpson E H. Measurement of diversity [J]. Nature, 1949, 163(4148): 688.
[20] 錢迎倩, 马克平. 生物多样性研究的原理与方法[M]. 北京: 中国科学技术出版社, 1994: 141-165.
[21] 何斌源, 范航清, 王 瑁, 等. 中国红树林湿地物种多样性及其形成[J]. 生态学报, 2007, 27(11): 4859-4870.
[22] 邱 霓, 徐颂军, 邱彭华, 等. 南沙湿地公园红树林物种多样性与空间分布格局[J]. 生态环境学报, 2017, 26(1): 27-35.
[23] 吴 瑞, 陈晓慧, 陈丹丹, 等. 海南省新英湾红树林资源现状调查分析[J]. 热带农业科学, 2016, 36(8): 35-37, 44.
[24] 林 生, 林惠宇. 海南东寨港国家级自然保护区及其周边陆域植物资源与植被调查研究[J]. 热带作物学报, 2018, 39(2): 398-404.
[25] 蔡开朗, 刘 俊, 曾德华. 三亚市铁炉港红树林自然保护区种子植物区系分析[J]. 湖南林业科技, 2018, 45(5): 73-77, 81.
[26] 张永夏, 张荣京, 邢福武, 等. 海南万宁青皮林群落多样性特征研究[J]. 西北植物学报, 2007(7): 1454-1460.
[27] 秦新生, 张荣京, 邢福武. 海南石灰岩地区的种子植物区系[J]. 华南农业大学学报, 2014, 35(3): 90-99.
[28] 张荣京, 秦新生, 陈红锋, 等. 海南俄贤岭石灰岩山地海南大戟灌丛群落研究[J]. 广西植物, 2007(5): 725-729.
[29] 贠建全, 邓双文, 陈红锋. 广东荷包岛维管束植物区系特征分析[J]. 植物科学学报, 2017, 35(1): 30-38.
[30] 莫耐波, 谢云珍, 覃康平, 等. 珍稀濒危植物瑶山苣苔伴生群落特征[J]. 广西林业科学, 2012, 41(3): 242-247.
[31] 宋贤利, 邢福武, 易绮婓, 等. 澳门松山阴香群落特征及物种多样性研究[J]. 福建林业科技, 2013, 40(3): 1-7.
[32] 中国植被编辑委员会. 中国植被[M]. 北京: 科学出版社, 1995: 402-405.
[33] 林 鹏. 中国红树林研究进展[J]. 厦门大学学报(自然科学版), 2001, 40(2): 592-603.
猜你喜欢
儿童故事画报·自然探秘(2022年3期)2022-04-27
发明与创新·小学生(2020年7期)2020-07-18
歌海(2018年4期)2018-05-14
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
科技创新导报(2016年21期)2016-12-17
现代经济信息(2016年13期)2016-06-17
少儿科学周刊·儿童版(2015年11期)2015-12-17
绿色科技(2015年6期)2015-08-05
学苑创造·A版(2014年10期)2014-10-20
