
短期封育对贵南县轻度退化高寒草甸植被特征的影响
2019-12-12 07:31贾志锋徐成体刘文辉魏小星雷生春
草业科学 2019年11期
贾志锋,马 祥,徐成体,刘文辉,魏小星,雷生春
(1. 青海大学畜牧兽医科学院,青海 西宁 810016;2. 青海省青藏高原优良牧草种质资源利用重点实验室 / 青海省畜牧兽医科学院,青海 西宁 810016)
草地是陆地上面积最大的生态系统之一,约占地球陆地面积的40%[1]。草地不仅是主要的畜牧业生产的基地,而且具有重要的生态服务功能[2-3]。然而,近几十年来,受自然因素和人为干扰等因素的影响,我国约90%的草地已经出现了不同程度的退化[4]。围栏封育是一种有效的退化草地恢复措施,以投资少、见效快、简单易行而被世界各国广泛应用[5-8]。
围封措施可以有效提高草地群落地上总生物量,明显增加草地植被盖度和牧草产量,逐步缓解草地沙化程度,有效控制土壤养分流失[9],改善草地土壤结构和营养资源状况,促进优质牧草生长[4-5,8,10]。赵新全等[11-12]指出,对于轻度退化高寒草地可以采用围栏封育进行治理。吉林省长岭县碱性退化草地进行围栏修复后发现,围栏区植被特征和土壤性质较放牧区显著提高,围栏后6年植被可以恢复[13]。研究表明,青藏高原东北缘高寒草甸地上生物量、地下生物量、总物种数、总密度和多样性指数随封育年限的延长呈先增加后降低的变化趋势,在封育6年后达最高;而长期围栏封育(> 9 年)却显著降低了青藏高原东北缘高寒草甸植物多样性、密度和生物量,致使群落结构单一化[14]。围栏封育有利于提高青藏高原退化高寒草地植物群落的盖度和生物量[15]。相同施肥水平下,围栏封育能显著提高藏北高寒草甸群落盖度,施加氮肥结合围栏封育对退化高寒草甸群落的恢复有促进作用[9]。
尽管已有大量有关高寒草甸围封的研究报道[8-15],但大多研究主要关注中长期围封后植被和土壤特征的变化,对短期封育的关注不够,同时,封育后草地植被特征的月动态和年动态变化尚不明确。为此,以青藏高原东缘轻度退化高寒草甸为研究对象,定量描述短期围栏封育对植被盖度、高度和功能群(禾本科、莎草科、豆科和杂类草)生物量的月动态和年动态的影响,以期为研究区及相似生态区草地修复治理提供科学依据。
1 试验材料与方法
1.1 试验地概况
试验地位于青藏高原东北缘的青海省贵南县森多乡茫什多滩(100.78° E,35.34° N,海拔3 562 m,图1),研究区气温低,日照长,辐射强,降水集中,雨热同季,但降水不足,蒸发量大,属典型的高原大陆性气候。年均气温2.6 ℃,年均降水量411.7 mm,6月-10月降水较多,牧草生长旺盛,2011年全年降水量530 mm,2012年降水量570 mm。年均日照时数2 907.8 h,年均蒸发量1 355.3 mm。草地类型为高寒草甸类,主要优势种有矮生嵩草(Kobresia humilis)、垂穗披碱草(Elymus nutans)、草(Koeleria cristata)和紫花针茅(Stipa capillata),毒杂草有黄花棘豆(Goxytropis ochrocephala)、狼毒(Stellera chamae)和高山唐松草(Thalictrum alpinum)。
1.2 试验设计及样品采集
依据草地植被和土壤特征,在研究区选取轻度退化高寒草甸为研究对象。研究区草地在生长季(5月-9月)自由放牧,放牧期内放牧强度约为7.9个羊单位·hm-2,非生长季(10月-次年4月)不放牧。放牧家畜为藏羊和牦牛。于2011年6月10日,采用随机区组试验设计,在研究区选取植被较为一致的草地建立3个面积分别为25 m2(5 m ×5 m)的围栏,相邻围栏间距为20 m,围栏自建立后全年禁牧。并以围栏外自由放牧草地为对照。放牧区和围栏区均进行鹰架(巢)建设,作为该试验的辅助措施。
于2011年7月-10月、2012年6月-10月的每月中旬,分别在围栏内外草地的对角线方向按“S”形随机设置6个面积为1 m2(1 m × 1 m)的样方进行植被盖度、高度、地上生物量的监测。首先采用针刺法测定植物分盖度,然后用钢卷尺测定植物自然高度,每种植物测定5株。最后将样方内的植物按照禾本科、莎草科和杂类草齐地剪下,分别装袋,带回实验室,在105 ℃杀青30 min,然后在70 ℃烘干至恒重,并称重。

图 1 研究区地理位置示意图Figure 1 Location of the study area
1.3 数据处理
采用Excel 2010对数据进行初步整理,采用SPSS 17.0 (SPSS 17.0, Chicago, IL, USA)软件独立样本T检验对围栏内外植被差异进行分析,同时,采用单因素方差分析(One-way ANOVA) Duncan和LSD法对不同月份间植被差异进行分析(P < 0.05)。数据结果用平均数 ± 标准差表示。采用Origin 2018作图。
2 结果与分析
2.1 短期围栏封育对群落盖度动态的影响
短期围栏封育措施对轻度退化草地盖度的影响较大(图2)。从年份间来看,2012年各封育处理草地盖度在7月-10月均高于2011年,分别高出34.6%、123.3%、43.3%和344.9%;放牧处理植被群落盖度在2012年也有小幅上升。封育或放牧处理下,不同取样时间群落盖度差异显著(P < 0.05)。
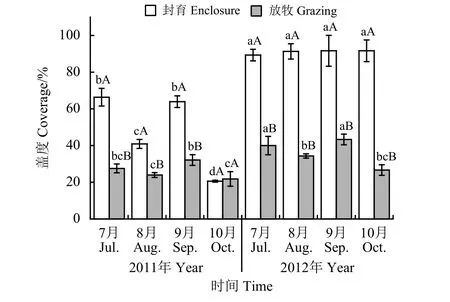
图 2 短期围栏封育对轻度退化高寒草甸植物群落盖度动态的影响Figure 2 Effects of short-term enclosure on the vegetation coverage of a lightly degraded alpine meadow
对不同处理来说,随着牧草生育期的延长,群落植被盖度的变化也有差异。2011年,封育处理植被盖度波动较大,7月份和9月份较高,8月份和10月份较低,而放牧组群落盖度整体变化较为平稳;2012年则不同,封育和放牧处理群落盖度均较平稳,波动不大。2011年,封育处理植被盖度在7月、8月、 9月均显著高于放牧处理(P <0.05),分别高出140.9%、70.9%和99.1%,但10月份(枯草期)与放牧处理相当(P > 0.05)。2012年,封育处理植被盖度在牧草生长期(7月-8月)、盛草期(9月)和枯草期(10月)均大幅提高,均显著高于放牧处理(P < 0.05),分别高出123.3%、166.0%、111.5%和243.8%。
2.2 短期围栏封育对群落高度动态的影响
不同处理对轻度退化草地群落高度的影响各异(图3)。对比两年的数据,可知,2012年封育处理各经济类群及群落高度均高于2011年,而放牧处理禾草、莎草类及群落高度较2011年无较大变化,杂类草植物高度则增加。不同处理各经济类群和群落高度在不同年份的波动也不同。封育或放牧处理下,不同取样时间间禾草类株高、莎草类株高、杂类草株高和群落高度差异显著(P <0.05)。2011年,随着牧草生育期的延长,封育处理禾草、杂类草和群落高度均呈现先升高后降低的单峰趋势,9月达到最高值,莎草类波动较大;放牧处理各经济类群和群落高度整体变化幅度较小,禾草类和群落株高逐渐降低,莎草和杂类草则为先升高后降低的趋势。2012年,7月-9月禾草类株高呈现出先升后降再升的规律,10月达到最大值;因禾草类株高值在群落内最大,群落株高受其影响较大,呈现出相同的趋势;莎草类株高整体变化不大;杂类草株高在7月达到最大值(11.2 cm),8月-10月基本无变化;放牧处理禾草、莎草类和群落高度总体均呈现逐渐降低的趋势,杂类草株高在8月达到最大值(8.2 cm),但整个生育期变化不大。
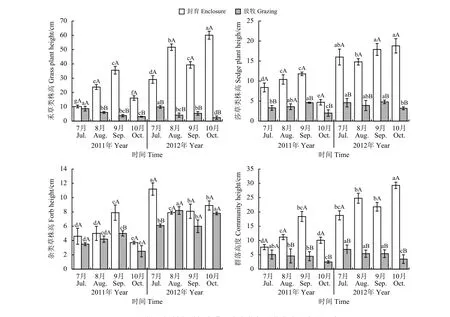
图 3 短期围栏封育对轻度退化高寒草甸群落高度动态的影响Figure 3 Effects of short-term enclosure on the vegetation height of a lightly degraded alpine meadow
与放牧相比,2011年封育处理在不同程度上提高了各经济类群和群落株高,但不同类群的响应有所差异。封育处理下禾草类和群落高度7月时较放牧处理无显著(P > 0.05)变化,8月-10月显著(P < 0.05)高于放牧处理;莎草类株高在7月、9月和10月显著高于放牧处理(P < 0.05),8月则无显著差异(P > 0.05);整个试验期内禾草、莎草和群落高度分别较放牧处理增加了294.9%、151.1%和183.8%。杂类草只在9月时显著高于放牧处理(P < 0.05),其他月份差异均不显著(P > 0.05)。2012年,封育处理对各经济类群的影响较2011年更大,整个试验期内禾草、莎草和群落高度均显著高于放牧处理(P < 0.05),分别较放牧处理增加722.8%、309.1%和346.7%。杂类草株高仅在7月时显著高于放牧处理(P < 0.05),而8月-10月均无显著差异(P > 0.05)。
2.3 短期围栏封育对地上生物量动态的影响
围栏封育对轻度退化草地地上生物量的影响较大(图4)。封育或放牧处理下,不同取样时间间禾本科地上生物量、莎草科地上生物量、杂类草地上生物量和总生物量差异显著(P < 0.05)。从年份上来看,封育处理禾草、莎草类和总生物量在2012年均高于2011年,7月-10月地上生物量的均值较2011年分别增加了12.4%、161.5%和52.8%,杂类草生物量降低了57.1%。可见,封育后次年莎草受到的影响更大;放牧处理下莎草和杂类草生物量变化不大,禾草类进一步降低,7月-10月的均值较2011年降低52.2%,总地上生物量也降低了20.9%。2011年,相较于放牧处理,封育可增加禾草、莎草、杂类草和总地上生物量,7月-10月的均值较放牧处理分别增加了210.9%、206.9%、50.7%和152.1%;2012年则不同,封育处理禾草、莎草类和总生物量7月-10月的均值较放牧处理分别增加了631.7%、540.3%和386.8%,增加幅度更大,且杂类草生物量7月-10月的均值较放牧处理下降了21.5%。
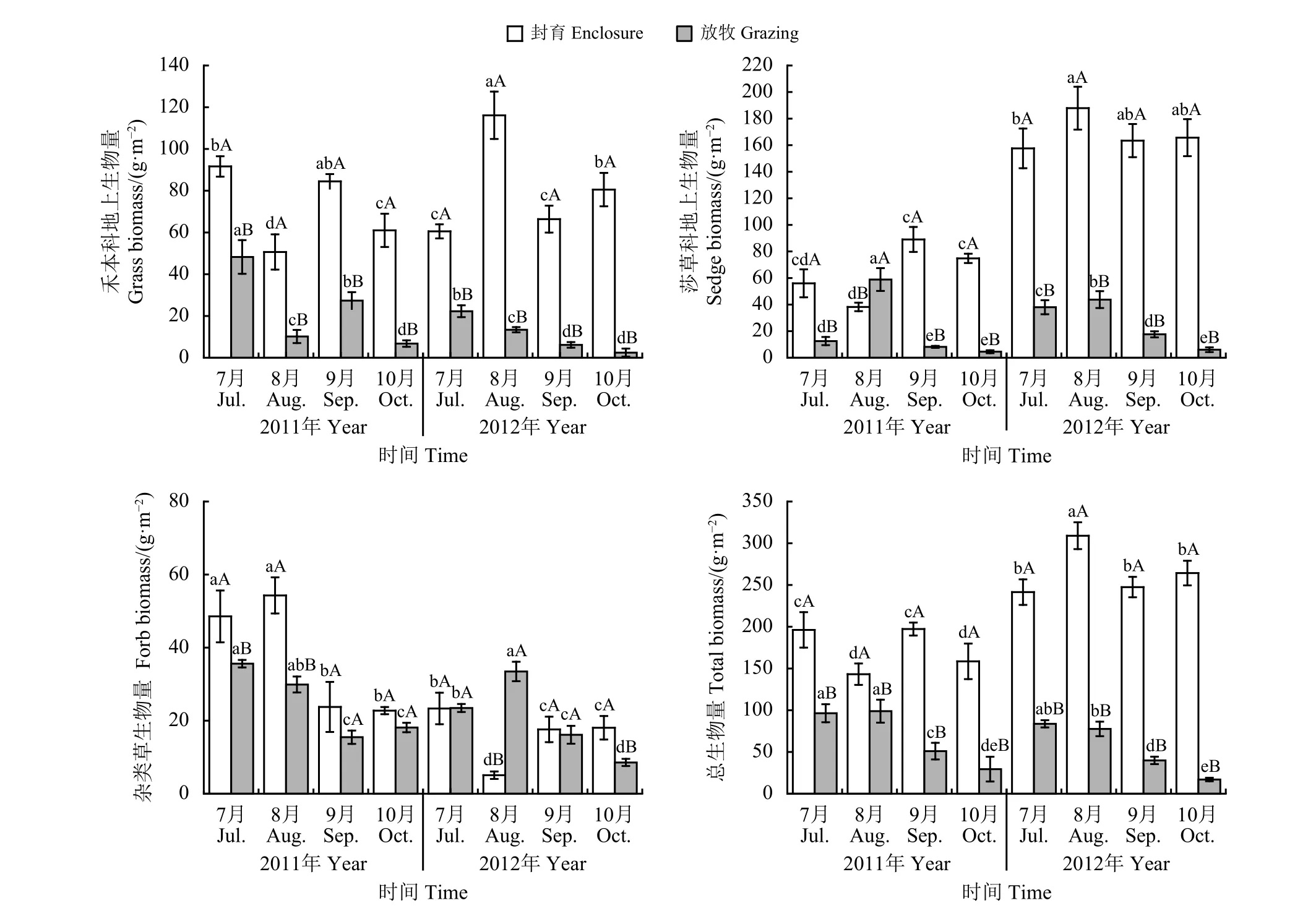
图 4 短期围栏封育对轻度退化高寒草甸地上生物量的影响Figure 4 Effects of short-term enclosure on the aboveground biomass of a lightly degraded alpine meadow
随着牧草生育期的延长,封育处理2011年禾草类和总地上生物量在7月-10月始终显著(P <0.05)高于放牧处理;莎草类生物量在7、9和10月显著(P < 0.05)高于放牧处理,但8月则显著(P < 0.05)低于放牧处理;杂类草生物量在7月 -8月显著高于放牧处理,9月-10月比放牧处理高,但差异不显著(P > 0.05)。2012年,封育处理禾草、莎草类和总地上生物量在7月-10月始终显著(P < 0.05)高于放牧处理,且增加幅度较大,7月-9月禾草类生物量较放牧处理分别增加了171.9%、766.7%、982.6%和3 218.1%,莎草生物量分别增加了314.7%、329.6%、828.8%和2 648.9%,地上总生物量分别增加了188.4%、298.3%、520.7%和1 453.1%;杂类草生物量在7和9月与放牧处理相差不大(P > 0.05),8月时显著(P < 0.05)显著低于放牧处理,10月则显著(P < 0.05)高于放牧处理。
2.4 短期围栏封育对草地各经济类群地上生物量比例的影响
短期围栏封育对草地地上生物量构成比例的影响如表1所列。可以看出,2011年封育处理地上生物量构成随着牧草生育期的延长而变化,7月 -8月禾草和杂类草生长迅速,7月时禾草占地上生物量比重最高,达46.7%;8月时杂类草为最高,达37.9%。9月-10月莎草生物量迅速增加,与禾草一起成为地上生物量的主要构成部分,占地上生物量的85%以上,杂类草则被抑制。放牧处理地上生物量构成波动较大,7月-9月禾草和莎草交替成为主要构成成分,杂类草生物量较稳定;10月时禾草和莎草生物量所占比重均降低,杂类草占到总地上生物量的61.6%。
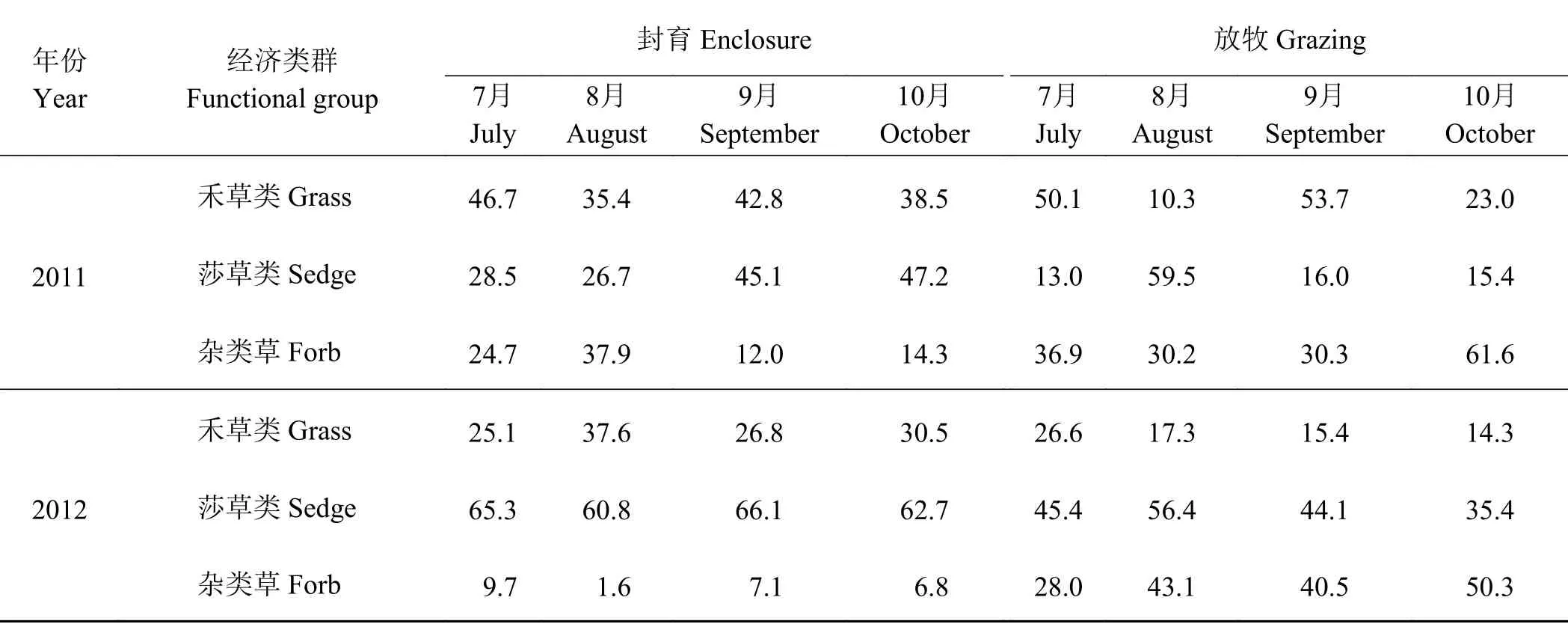
表 1 短期封育处理对各类群地上生物量构成比例的影响Table 1 Effects of short-term enclosure on the composition of the aboveground biomass%
2012年则不同,封育处理各经济类群生物量占总地上生物量的比重较为稳定,7月-10月均以莎草类为主要构成部分,占60%以上,杂类草被抑制在10%以下,禾草保持在30%左右。放牧处理下禾草生物量所占比重较低,莎草和杂类草成为地上生物量的主要构成部分,且杂类草的比重呈上升趋势,10月时占到50.3%。
3 讨论
草地生物量不仅是群落功能和结果的测度指标,还是生态系统功能和结构的基础,同时生物量可以反映退化草地的恢复效果[16-17]。本研究发现,短期围栏封育可以提高轻度退化草地的高度、盖度和地上生物量,其中封育第2年10月时植被盖度高于放牧243.8%、总生物量较放牧处理高1 453.1%。对小嵩草(Kobrecia parva)高寒草甸3年的封育研究表明,封育处理后轻度退化草地的总盖度以及禾本科牧草的分盖度有了明显提高[17];宋梅玲等[18]研究了包括禁牧(禁)在内的5种草地管理方式对狼毒型退化草地的草产量的影响,结果表明,禁牧封育处理下禾本科植物的生物量与总生物量显著高于对照;研究发现,在高寒草甸草地内,禁牧封育第2年与禁牧封育当年相比,莎草科牧草、禾本科牧草产量分别增加3.3%和5.1%,杂类草由50.9%下降到42.5%,鲜草总产量增加320 kg·hm-2,杂类草在草群中的比重降低,优良牧草在草群中的比重升高[19];退化草地改良试验研究表明,围栏封育使禾本科牧草和豆科牧草分别增加26.81%~39.85%和20.74%~37.85%,使莎草科牧草比重降低11.85%~19.23%[20]。本研究结果与前人研究结果相似[17-20],是可行的草地退化治理措施。
草地不同经济类群有其自身生长发育的特点,在环境变化时呈现具有其自身特性的变化,即各经济类群在封育后呈现不同的变化趋势。在本研究中,封育次年草地群落7月-10月均以莎草类为主要构成部分,占60%以上,杂类草被抑制在10%以下,禾草保持在30%左右,其原因是莎草类植物的物候期较早,返青时间更早,对水肥等资源的利用在时间上比其他植物更提前,能在禾草、杂类草等植物密闭之前提早利用有效资源,以满足其生长发育的要求[21]。本研究中围封后次年莎草生物量一直保持在60%以上,群落结构变化较小。同时,繁茂的莎草和禾草也抑制了杂类草的生长,杂类草生物量比围封初期明显降低。同一种植物类群对围封的响应在年际间也有差别。围栏封育初期,禾草和杂类草生长迅速,7月时禾草占地上生物量比重最高(46.7%),8月时杂类草为最高(37.9%),9月-10月及次年整个试验期莎草生物量在总地上生物量所占比重上升,次年一直保持在60%以上,而杂类草则被抑制在10%以下。围栏封育通过人为消除家畜对草地生态系统的影响,使其依靠自身的弹性得以恢复和重建[22]。围封初期,对比放牧处理可知禾草和杂类草为优势物种,生物量较大,放牧压力的消失使得放牧过程中占优势的禾草和杂类草植物得以迅速生长恢复,在一定程度上抑制了莎草类植物的生长。而经过一段时间的围封后(9月-10月),消除了高寒草甸生态系统的养分限制,使得养分利用效率高的莎草植物迅速生长,增加了植被高度和盖度,使得群落结构和植物竞争格局发生改变,从地下养分竞争转为地上光和空间竞争[23]。因此,围封后总地上生物量的增加不仅与放牧压力的降低有关,还受围封后土壤性质改善的影响。
关于围栏封育时间,研究表明禁牧封育时间并非越长越好,长期围栏封育并不能增加草地群落的生产力及其放牧功能[24]。封育时间过长,会使草地中枯草大量堆积,造成凋落物的大量积累,抑制植物的再生和幼苗的形成,不利于草地的繁殖更新[25],并且高大杂草丛生,影响草地资源的牧用价值[19],进而影响草地的再生功能[18]。本研究中采用短期围栏措施,且由于降水量等自然因素的影响,放牧区的群落盖度在2011年与2012年间有所差异,与2011年相比,2012年草地盖度整体更高;同时封育处理在两年间也有显著的差异,特别是在8月和10月,2012年封育处理草地盖度在8月和10月分别较2011年高出123.3%和344.9%。放牧对照的群落高度在2011年与2012年间差异不明显;但封育处理在两年间有显著的差异,除9月外,其他月份的草地高度均呈现2012年显著高于2011年的趋势。短期封育处理的总地上生物量表现出与群落盖度相似的趋势。短期封育处理可以明显提高退化草地的高度、盖度和地上生物量,是该区域轻度退化草地恢复的有效措施。
本研究发现,短期围栏封育对轻度退化草地的恢复是多个因素的综合效果,首先,短期围栏封育处理首先减少了放牧家畜和野生动物对草地的采食和践踏,草地植物的生长受到的影响大幅度减弱;其次,短期封育对草地种子库有影响,由于短期封育处理草能完成营养生长和生殖生长的整个生育过程,能产生种子,对于种子库的大小具有重要意义;最后,鹰架(巢)架设后鹰的种类和数量增加,害鼠种群数量由建设前的690个·hm-2减少到243个·hm-2,降低了64.78%,降低了对草地的破坏,也可能促进了轻度退化草地牧草的生长。多个因素的综合结果使得短期围栏封育草地的牧草个体的生长、群落结构和生物量发生改变。
4 结论
短期围栏封育可显著增加贵南县轻度退化草地植被群落的盖度、高度和地上总生物量,且不同经济类群对围封的响应在时间上有所不同,围封初期禾草和杂类草植物迅速生长,莎草则在9月后逐渐占据优势。围封次年莎草生物量占地上总生物量的60%以上,禾草占30%左右,杂类草被抑制在10%以下。因此,短期围栏封育措施可用于贵南县轻度退化草地的恢复管理。
猜你喜欢
疯狂英语·新读写(2022年1期)2022-11-23
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
疯狂英语·读写版(2022年1期)2022-01-29
绿色科技(2021年10期)2021-06-23
文史博览·文史(2021年1期)2021-04-01
林业科技(2020年3期)2021-01-21
畜牧兽医科学(2019年24期)2019-09-10
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
