
内生真菌诱导子通过MeJA介导油樟细胞挥发油积累的研究
2019-12-05 01:25:50严宽,刘杰,魏琴,谢浩
福建茶叶 2019年9期
严 宽,刘 杰,魏 琴,谢 浩
(1.宜宾学院川茶学院;2.宜宾学院生命科学与食品工程学院,宜宾 644000)
油樟[Cinnamomum longepaniculatum(Gamble)N.Chao]主要分布于四川省,其茎、叶等器官可以炼取大量精油,主要为萜类化合物,包含了1,8-桉叶油素、α-松油醇、γ-松油烯等化学物质。油樟精油具有一定的功效性,包括消炎、杀菌和抗氧化等作用。植物内生真菌诱导子诱导植物的次生代谢是通过特定的信号分子和其相应的信号转导途径实现的。随着对信号分子和信号转导途径的深入研究,发现在许多植物体中MeJA都发挥着重要的信号分子作用[1]。在施加外源MeJA时,可以诱导植物产生防御机制,诱发多种次生代谢产物的生成[2]。因此,本实验通过对内生真菌诱导子促进油樟悬浮细胞产生挥发油合成过程中MeJA作用的研究,来探讨内生真菌诱导子与油樟悬浮细胞挥发油合成过程中的MeJA信号的转导机制,以便在未来关于油樟细胞中次级代谢的调控规律能够有更深入的了解,从而可以找到提高调控油樟细胞挥发油产量的方法。
1 方法与材料
1.1 材料来源与处理
从宜宾红岩山油樟基地(位于27°50'N;105°20'E)采集了油樟叶的样本。内生真菌(Penicillium commune,2J1)从油樟植物中分离出来,将这些真菌培养并保存在PDA培养基内。
1.2 建立油樟细胞悬浮体系
选用生长良好的油樟幼嫩叶片培养愈伤组织后,继代培养2-3次。之后选取色泽鲜亮、长势良好,质地疏松的嫩黄色愈伤组织,约2.0g/瓶,25℃,遮光,振荡培养(120 r·min-1)。每隔7 d继代一次,连续继代2-3次[3]。
1.3 内生真菌诱导子的制备
将一株青霉菌属(Penicilliumsp.)的内生真菌接种到配置好的PDA液体培养基中,在28℃,130r/min的震荡培养箱中培养7d,之后将2J1的菌体与发酵液进行分离,将内生真菌2J1的菌体破碎成匀浆之后与发酵液混合,进行减压抽滤,最后在高温高压灭菌锅中121℃灭菌20min,便可得到内生真菌诱导子。采用蒽酮-硫酸法测定诱导子糖含量[4]。
1.4 油樟悬浮细胞挥发油的提取与测定
精确称取烘干之后的油樟细胞0.3g,加入环己烷冷浸过夜(环己烷与细胞的比例为4:1),随后进行30min的超声提取,在5000r/min的超速离心机上离心4min,提取上清液,定容到5mL。用针筒抽取液体,滤膜过滤至上样瓶中,测定挥发油的含量,用GC-MS分析[5]。1,8-桉叶油素的标准曲线Y=73900X-299200,R2=0.9993。
1.5 油樟悬浮细胞中MeJA含量的测定
油樟悬浮细胞中MeJA含量采用顶空固相微萃取技术测定[14]。标准曲线:y=7676.9x+6234.5 R2=0.9985
1.6 数据软件及处理
挥发油的产量和实验处理使用三次重复测量。所有相关数据采用SPSS进行路径系数分析统计19.0和Excel2007进行数据分析。
2 结果与分析
2.1 内生真菌诱导子对油樟悬浮细胞MeJA释放量和挥发油积累的影响
在培养7d的油樟悬浮细胞中加入内生真菌2J1粗诱导子40mg/L,以加入等量的PDA培养基为对照组。考察2J1诱导子对油樟悬浮细胞内MeJA含量和挥发油积累的影响。经过油樟内生真菌2J1处理后的油樟悬浮细胞,培养至第21d时诱导效果最好,产生的挥发油含量较高;但是诱导的时间过长,油樟悬浮细胞挥发油产量反而有所减少。
2.2 MeJA释放和挥发油积累与不同油樟内生真菌诱导子浓度的关系
分别向培养了7d的油樟悬浮细胞中加入浓度为0mg/L、20mg/L、40 mg/L、60mg/L、80mg/L的内生真菌2J1粗诱导子,在油樟悬浮细胞分别培养21h和14d后检测不同浓度诱导子对油樟悬浮细胞产生的MeJA释放和挥发油的积累的影响。
2.3 MeJA对内生真菌诱导子促进油樟悬浮细胞挥发油合成的影响
为了更好的了解内生真菌2J1粗诱导子通过化学信号MeJA诱导促进油樟悬浮细胞产生挥发油的作用机制,在油樟悬浮细胞培养到7d后进行了如下处理,设置了如下图中的对照试验,通过检测其中各组中油樟悬浮细胞MeJA(图1)和挥发油的合成情况(图2)进行分析研究,其中内生真菌2J1粗诱导子、SHAM、外源MeJA的添加浓度分别为40 mg/L、2mmol/L、100μmol/L。检测油樟悬浮细胞MeJA释放量的时间为处理后21h,检测油樟悬浮细胞挥发油的合成时间为处理后14d。
结果表明,添加外源MeJA可以促使油樟细胞中的挥发油合成。而SHAM则可抑制细胞中MeJA的积累,并导致挥发油产量的降低。但是,添加SHAM没有使内生真菌诱导子诱导的油樟细胞挥发油的合成量削减完全,因此在油樟悬浮细胞中可能存在由MeJA作为信号分子的调控挥发油合成的信号途径。
3 讨论
由于内生真菌诱导子属于细胞外的物质不能直接进入细胞发挥作用,所以需要依靠植物细胞膜外的特异性受体,当它与细胞膜上受体结合后,便会使细胞的结构发生变化,产生出一些特定的胞内信使物质,这些信使物质可以通过一系列的信号转导进而实现对特定基因的调控作用,使植物细胞中的防御性次生代谢系得以激活,进而可以使植物次生代谢产物合成增加[6]。因此,MeJA的迸发可能是经2J1粗诱导子处理后出现的前期反应,MeJA可能属于胞内的一种信使物质,经过一系列复杂的生化反应,成功诱导了油樟细胞中挥发油产量的提升。本研究虽然对MeJA介导内生真菌诱导子对油樟挥发油合成的调控机制做了进一步研究。因此为了能够使诱导子充分的发挥其诱导作用,关于内生真菌2J1粗诱导子的纯度和结构有必要做进一步的纯化和检测,以此来研究诱导子更好的制备和保存方法[7]。由此实验还可以推测出存在着其他途径引起油樟细胞中挥发油的合成与积累。植物次生代谢产物的合成途径是非常复杂的,可能还存在其他的信号转导途径,这仍需进一步的研究。
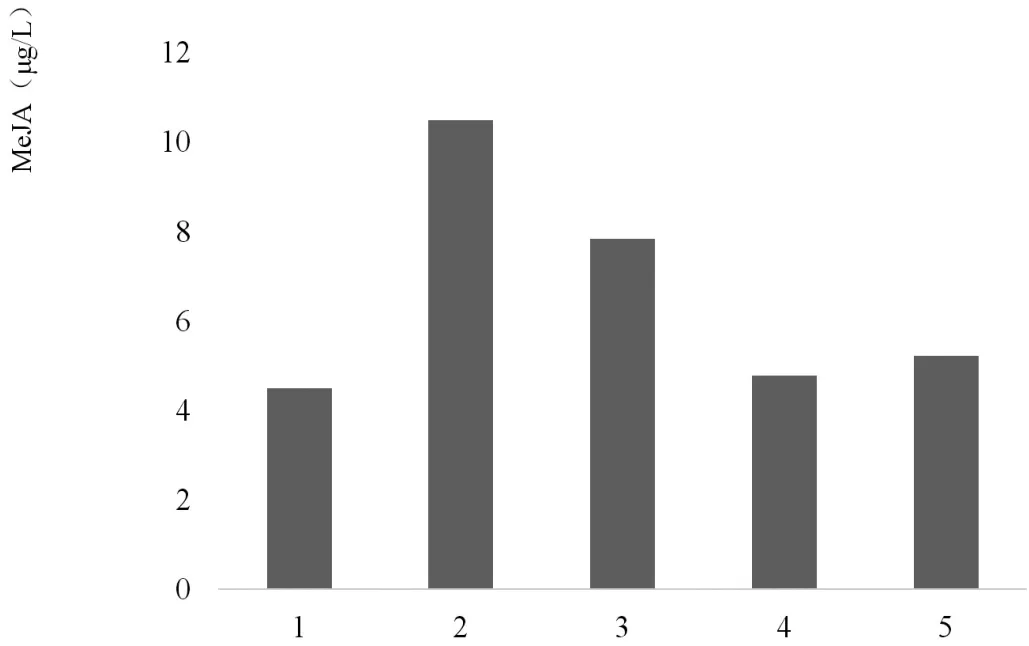
图1 各组中MeJA的累积情况
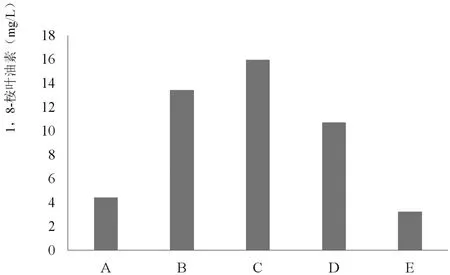
图2 各组中挥发油的累积情况
猜你喜欢
绿色天府(2022年2期)2022-03-16 07:06:12
广州化工(2021年22期)2021-11-30 13:21:10
云南医药(2021年3期)2021-07-21 05:40:20
湖北农机化(2020年4期)2020-07-24 09:07:16
世界农药(2019年4期)2019-12-30 06:25:10
今日农业(2019年11期)2019-08-15 00:56:32
乡村地理(2018年2期)2018-09-19 06:44:02
四川林业科技(2015年6期)2015-12-02 04:18:38
中国医药导报(2015年24期)2015-02-28 22:07:30
宜宾学院学报(2014年12期)2014-07-20 11:54:15
