
精胺对鹅免疫器官指数及免疫相关因子基因表达的影响
2019-12-04 08:05易治鑫蒋易龙王秋宏徐麒麟王新兴莫桂林姜冬梅康波
浙江大学学报(农业与生命科学版) 2019年5期
易治鑫,蒋易龙,王秋宏,徐麒麟,王新兴,莫桂林,姜冬梅,康波*
(1.四川农业大学动物科技学院,成都611130;2.巴中市农林科学研究院,四川 巴中636000;3.成都市农林科学院,成都611130)
多胺(主要包括腐胺、亚精胺和精胺)能作为内源性免疫调节因子参与调节机体免疫[1-2]。研究发现,精胺作为内源性细胞因子抑制剂,能抑制白细胞介素-1(interleukin-1, IL-1)、白细胞介素-6(interleukin-6, IL-6)及肿瘤坏死因子α(tumor necrosis factor-α, TNF-α)的合成[3-5]。HASKÓ 等[1]研究发现,精胺不仅能抑制脾细胞释放γ-干扰素(interferon-γ,IFN-γ),还能抑制巨噬细胞翻译白细胞介素-12(interleukin-12, IL-12),进而发挥其免疫调节作用。CAO 等[6]通过在仔猪日粮中添加0.4 mmol/kg 精胺发现,精胺不仅能显著抑制胸腺和脾中TNF-α、IL-1β、IL-6 和IFN-γ 基因表达,同时还能显著提高IgM、IL-10 及转化生长因子β1 基因的表达。该结果提示精胺能介导免疫相关因子基因的表达,从而影响机体正常生理状态下的免疫功能。此外,当母乳中精胺浓度低于2 nmol/L 时,80%的新生儿会出现过敏反应;而精胺浓度在13 nmol/L 以上时,过敏症状发生概率将显著降低[7]。这说明精胺对幼龄动物的免疫能力具有调控作用。综上所述,精胺能介导免疫因子参与的机体免疫调控。
胸腺、脾和法氏囊作为禽类重要的免疫器官,是产生免疫细胞和免疫因子的主要场所[8];肝不仅能参与代谢和解毒,在禽类免疫功能中也具有十分重要的作用。我们的前期研究表明,精胺可通过介导鹅卵泡颗粒细胞雌激素受体和促黄体激素受体基因表达来参与调控雌鹅卵巢功能[9-10]。然而,目前尚未见精胺对雌鹅免疫器官指数和免疫相关因子基因表达影响的研究报道。因此,本研究用不同浓度精胺灌喂四川白鹅雌鹅2 周后,测定肝、胸腺、脾和法氏囊的免疫器官指数,并定量检测肝、胸腺、脾和法氏囊组织中TNF-α、IL-1β、IL-6 和IFN-γ 基因的表达量,以明确外源性精胺对雌鹅免疫器官指数和免疫相关因子基因表达的影响,为探究精胺调控家禽免疫功能的作用机制提供参考。
1 材料与方法
1.1 实验动物及样品采集
随机选取四川农业大学家禽育种实验场同批孵化、相同环境饲养的成年四川白鹅母鹅18 只,随机分为3组(n=6),自由采食和饮水。对照组灌喂禽用生理盐水,试验组根据鹅体质量分别灌喂5 mg/kg和10 mg/kg 精胺,每日于8:00 和20:00 进行灌喂。灌喂2周后,于颈部放血处死,迅速采集鹅肝、胸腺、脾和法氏囊组织,称量,用于后续试验。所有实验操作都严格按照四川农业大学实验动物操作规范与福利管理委员会制定的《实验动物操作规范》(川农大校发〔2014〕18号)进行。
1.2 免疫器官指数测算
鹅肝、胸腺、脾和法氏囊组织的器官指数计算公式如下:免疫器官指数/(g/kg)=免疫器官鲜质量/g÷活体质量/kg。
1.3 组织总RNA 提取和cDNA 制备
参照Trizol 试剂(TaKaRa 公司,大连)操作说明书提取组织样品总RNA,并取3 μL 总RNA 通过1.5%琼脂糖凝胶电泳检测总RNA质量。参照反转录试剂盒(TaKaRa 公司,大连)操作说明书制备cDNA。
1.4 实时荧光定量聚合酶链式反应
实时荧光定量聚合酶链式反应(polymerase chain reaction,PCR)体系:iTaqTM通用型SYBR Green超预混液(Bio-Rad Laboratories, CA, USA)5 μL,上游和下游引物各0.2 μL,cDNA 模板0.5 μL,无RNA酶水4.1 μL,共10 μL。反应条件:95 ℃预变性3 min;95 ℃变性10 s,根据引物退火温度(表1)分别退火30 s,72 ℃延伸30 s并采集荧光,共45个循环。绘制熔解曲线。以甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH)作为内参基因,采用2-ΔΔCT方法计算目的基因的相对表达量,每个样品设3个重复。
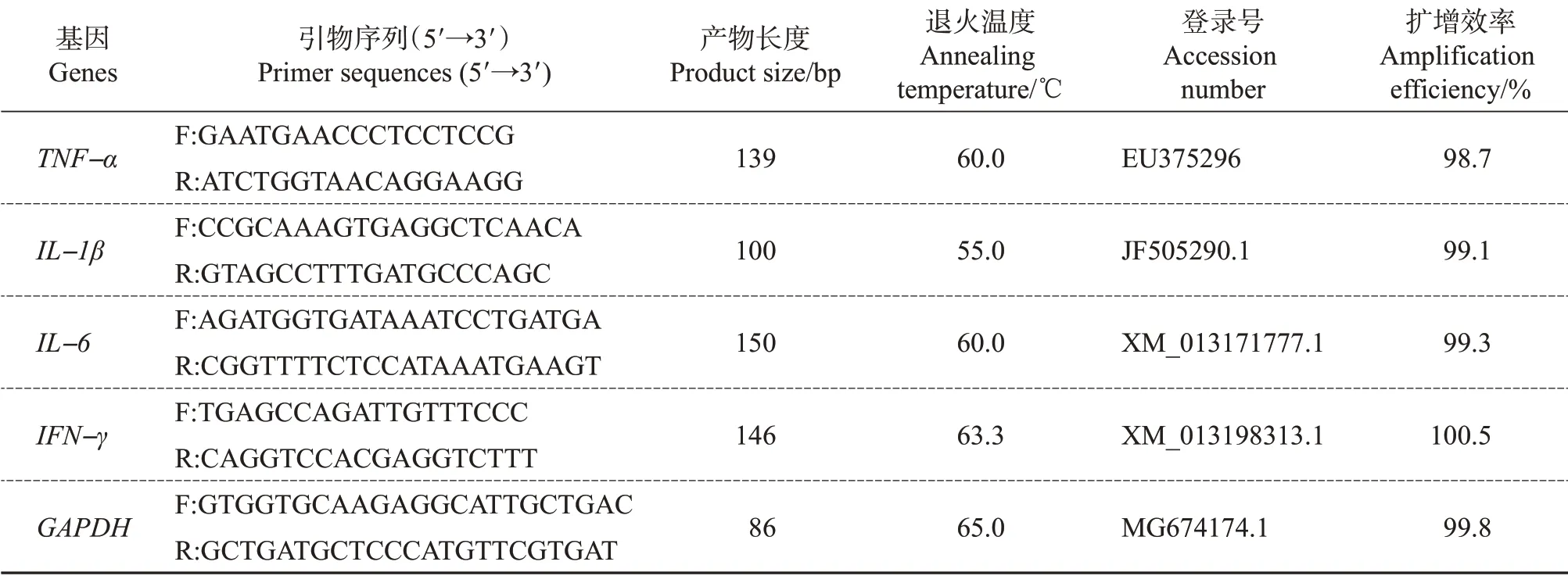
表1 引物序列信息Table 1 List of primer sequences
1.5 免疫器官组织多胺含量检测
参照KANG 等[10]报道的方法,采用苯甲酰氯柱前衍生并联合高效液相色谱检测鹅肝、胸腺、脾和法氏囊组织中的多胺含量。
1.6 数据统计与分析
应用SAS 9.2统计分析软件的均值过程进行描述性统计,并进行方差分析和邓肯多重比较。数据用平均值±标准误表示,P<0.05表示差异有统计学意义。
2 结果与分析
2.1 鹅免疫器官组织RNA 提取及免疫相关因子基因扩增结果
如图1 所示,鹅肝、胸腺、脾和法氏囊组织总RNA的28S和18S片段条带清晰,未产生降解的5S片段,表明肝、胸腺、脾和法氏囊组织总RNA质量良好,可用于后续试验。
如图2 所示,利用已提取的鹅免疫器官组织总RNA(以肝为例),对TNF-α、IL-1β、IL-6、IFN-γ 和GAPDH基因的特异性引物进行PCR扩增的结果表明,目的基因片段条带单一,大小与预期片段相一致;此外,如图3所示,TNF-α、IL-1β、IL-6、IFN-γ和GAPDH基因实时荧光定量PCR熔解曲线呈现单一峰型,表明引物特异性高。因此,本文所设计的免疫因子基因引物可用于后续实时荧光定量PCR试验。
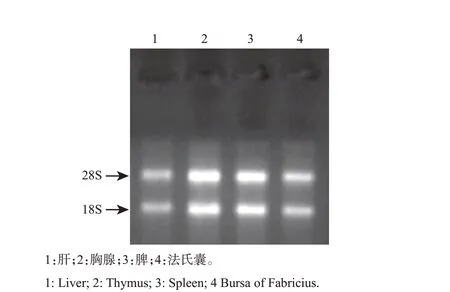
图1 鹅免疫器官组织总RNA电泳图Fig.1 Electrophoretogram of total RNA in the immune organ of female geese

图2 鹅肝组织目的基因RT-PCR产物电泳图Fig.2 Electrophoretogram of RT-PCR products for target genes in the liver of female geese
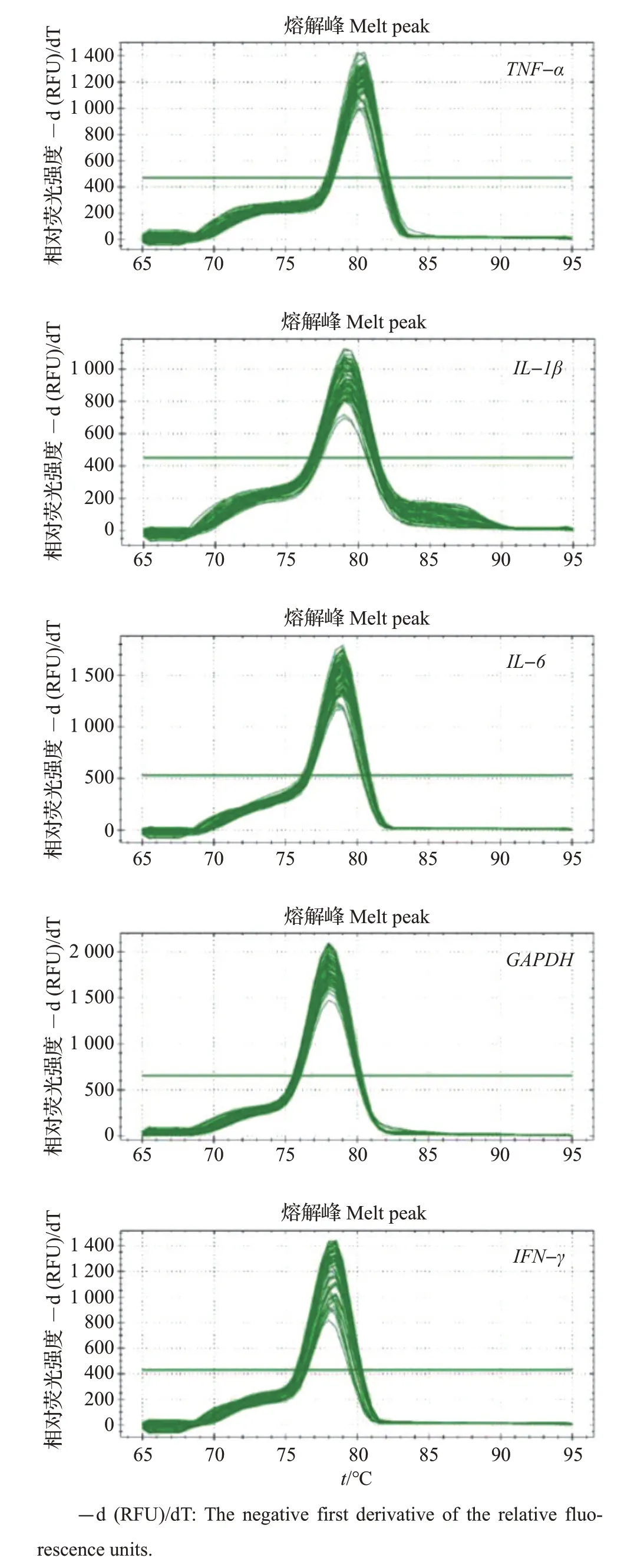
图3 目的基因实时荧光定量PCR熔解曲线Fig.3 Melt curves of real-time quantitative PCR amplifications of target genes
2.2 精胺对雌鹅免疫器官指数的影响
如图4所示:5 mg/kg精胺灌喂组肝指数与对照组无显著差异(P>0.05),而10 mg/kg精胺灌喂组鹅肝指数显著低于对照组(P<0.05);不同浓度精胺处理对雌鹅胸腺、脾和法氏囊指数均无显著影响(P>0.05)。

图4 精胺对鹅免疫器官指数的影响Fig.4 Effects of spermine on immune organ indexes of the liver,thymus,spleen and bursa of Fabricius in female geese
2.3 精胺对鹅免疫器官免疫相关因子基因表达的影响
由图5 可知:不同浓度精胺灌喂对鹅肝组织TNF-α、IL-1β 和IL-6 基因表达较对照无显著影响(P>0.05),在10 mg/kg 精胺灌喂组的鹅肝组织中IFN-γ 表达量显著低于对照组,是对照组的39.37%(P<0.05)。5 mg/kg 精胺灌喂后,鹅胸腺组织中TNF-α、IL-1β、IL-6 和IFN-γ 基因表达量均显著低于对照组(P<0.05),分别是对照组的40.00%、45.88%、45.30%和11.00%,10 mg/kg 精胺处理对鹅胸腺TNF-α、IL-1β、IL-6 和IFN-γ 基因表达无显著影响(P>0.05)。5 mg/kg 和10 mg/kg精胺灌喂后,鹅脾组织IFN-γ 基因表达量分别是对照组的46.45%和39.04%(P<0.05),而对脾组织TNF-α、IL-1β 和IL-6 基因表达均无显著影响(P>0.05)。5 mg/kg精胺处理组的鹅法氏囊TNF-α、IL-1β、IL-6 和IFN-γ 基因表达量分别是对照组的32.84%、49.46%、27.89% 和19.97%(P<0.05);10 mg/kg精胺处理组的法氏囊TNF-α和IFN-γ基因表达量分别是对照组的78.51%和81.82%(P<0.05)。
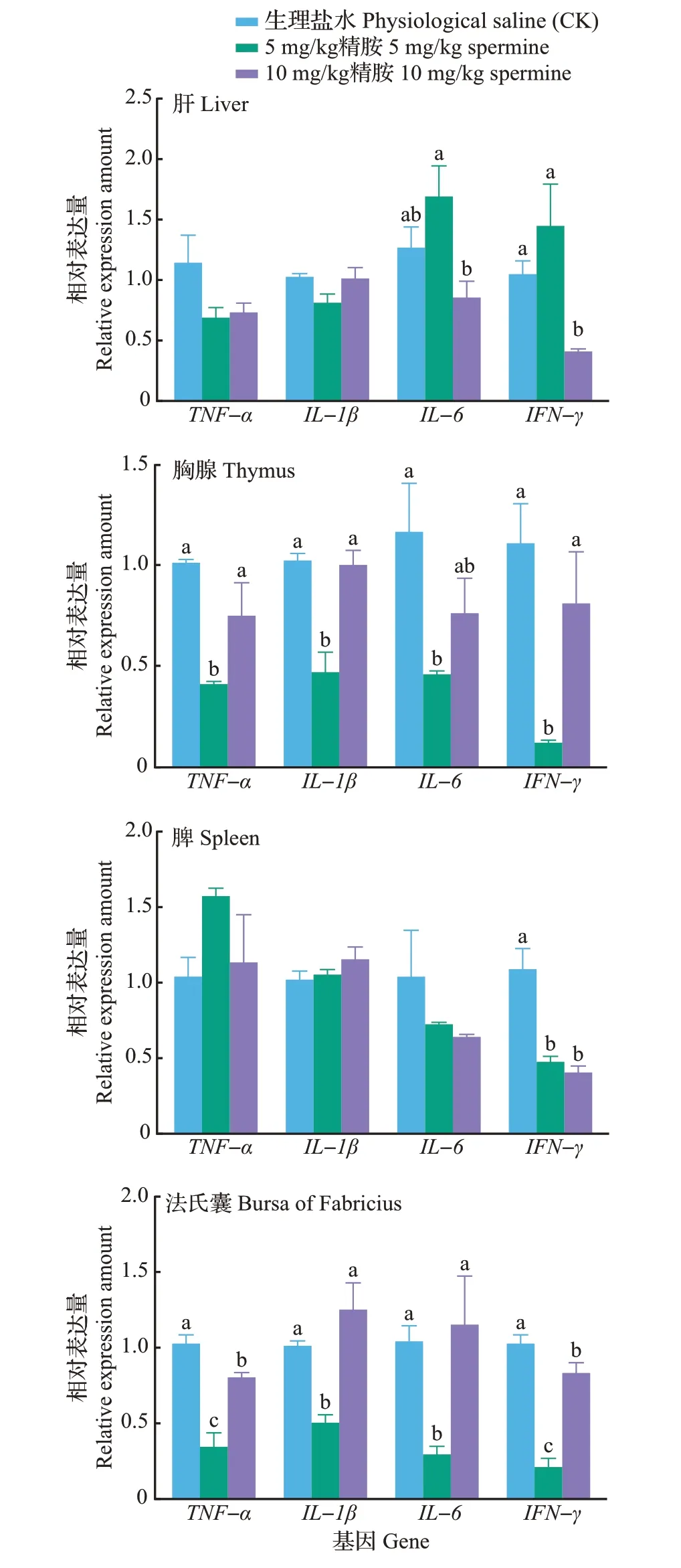
图5 精胺对鹅免疫相关因子基因表达的影响Fig.5 Effects of spermine on expression levels of genes related to immune factors in female geese
2.4 精胺对鹅免疫器官多胺代谢的影响
如图6 所示:外源性精胺处理显著提高了鹅肝组织精胺含量(P<0.05),而对腐胺和亚精胺含量均无显著影响(P>0.05);10 mg/kg精胺处理显著增加了法氏囊组织腐胺含量(P<0.05),显著降低了精胺含量(P<0.05),而对亚精胺水平无显著影响(P>0.05);不同浓度精胺对鹅脾和胸腺组织中腐胺、亚精胺和精胺含量均无显著影响(P>0.05)。
3 讨论与结论
多胺可作为内源性免疫调节因子,在调控机体免疫功能中发挥着重要作用[11-12]。本实验室人员前期研究发现,在正常生理条件下低浓度亚精胺能抑制小鼠TNF-α 和IL-1β 基因的表达,而高浓度亚精胺则会促进IL-6和IFN-γ基因的表达,提示免疫相关因子表达对亚精胺存在剂量依赖性[13]。OKUMURA 等[2]研究发现,多胺能显著降低大鼠肝缺血再灌注后血清中IL-1β、IL-6、TNF-α 和IFN-γ的水平,对趋化因子也有显著抑制作用。此外,研究表明,在小胶质细胞和急性胰腺炎中,多胺具有抑制TNF-α和IL-6基因表达的作用[14-15]。
胸腺、法氏囊和脾是禽类主要的免疫器官[16],而胸腺指数、法氏囊指数和脾指数能够反映机体免疫水平,在判断禽类免疫功能中具有重要意义[17]。研究发现,炎症、致敏及感染组织中精胺含量显著升高,提示精胺在炎症和致敏等病理过程中发挥着重要作用[5,18]。本研究结果表明:不同浓度精胺灌喂对鹅胸腺指数、脾指数和法氏囊指数无显著影响,在脾和胸腺组织中腐胺、亚精胺和精胺含量均无显著变化;10 mg/kg精胺处理显著下调鹅肝指数,同时,肝组织中精胺水平显著升高,提示外源性精胺在不同组织中沉积具有组织特异性,肝组织对外源性精胺处理更加敏感。另外,10 mg/kg精胺处理后,鹅法氏囊组织精胺水平显著降低,而腐胺水平显著增加,提示外源性精胺可能加速了法氏囊组织中的多胺分解代谢,促使精胺向腐胺转化。这进一步说明不同组织中的多胺代谢能力及其对精胺的耐受量可能存在差异。

图6 精胺对鹅免疫器官组织多胺水平的影响Fig.6 Effects of spermine on polyamine contents in the liver,thymus,spleen and bursa of Fabricius of female geese
精胺能通过发挥其免疫调节因子功能参与调控机体的免疫功能。ZHANG等[19-20]研究发现,精胺能抑制脂多糖诱导的人单核细胞TNF-α的释放,提示精胺具有参与调控人免疫功能的潜力。HASKÓ等[1]也发现,精胺能通过介导巨噬细胞IL 的分泌来参与调节机体免疫。高水平TNF-α、IL-1β 和IL-6能诱导组织发生炎症反应,而精胺在转录水平上能通过抑制TNF-α、IL-1β 和IL-6 基因表达进而参与调节机体免疫功能[2,21-22]。本研究发现,5 mg/kg精胺处理后,鹅胸腺和法氏囊组织中TNF-α、IL-1β 和IL-6 基因表达量均显著降低,与上述研究结果相似,进一步说明精胺可介导胸腺和法氏囊组织中TNF-α、IL-1β和IL-6基因表达,进而在转录水平上调控机体免疫功能。同时,10 mg/kg 精胺对胸腺和法氏囊组织中IL-1β 和IL-6 表达无显著影响,提示精胺介导胸腺和法氏囊中IL-1β和IL-6的表达可能具有剂量依赖性。此外,外源性精胺处理后,鹅肝和脾组织中TNF-α、IL-1β 和IL-6 基因表达量相较于对照组并未发生显著变化,提示精胺对TNF-α、IL-1β和IL-6基因表达的调控可能具有组织特异性。
IFN-γ是由T细胞、B细胞和自然杀伤细胞等分泌的Ⅱ型干扰素[23],可通过激活效应免疫细胞、促进抗原呈递进而发挥其抗菌、抗病毒和抗肿瘤作用[24-25]。有研究发现,精胺对脾组织中IFN-γ合成释放有抑制作用,进而调控机体免疫能力[1]。在日粮中添加精胺能抑制仔猪胸腺和脾组织中IFN-γ基因表达[6]。本研究结果表明,外源性精胺显著抑制鹅脾中IFN-γ基因表达,5 mg/kg精胺可抑制胸腺和法氏囊中IFN-γ基因表达,这与上述结果相似,说明精胺不仅能通过翻译水平调控脾组织中IFN-γ 合成,还能在转录水平上影响脾组织中IFN-γ基因的表达[1,6]。
总之,本研究结果表明,精胺可通过介导雌鹅免疫相关因子TNF-α、IL-1β、IL-6 和IFN-γ 基因表达来参与调控鹅免疫功能,并存在剂量依赖性和组织特异性。
猜你喜欢
吉林畜牧兽医(2022年2期)2022-11-16
河南科技学院学报(自然科学版)(2022年1期)2022-02-25
中国农业大学学报(2022年2期)2022-01-05
郑州大学学报(医学版)(2021年6期)2021-12-14
流行色(2021年8期)2021-11-09
国际医学放射学杂志(2021年4期)2021-08-05
皮肤病与性病(2021年3期)2021-07-30
汉语世界(The World of Chinese)(2019年5期)2019-11-11
少儿科学周刊·少年版(2018年12期)2018-01-26
科学中国人(2017年36期)2017-06-09
