
臭氧处理对交链孢菌生长及其毒素积累的抑制作用
2019-12-03 01:08王刘庆李祖梁
食品科学 2019年21期
姜 楠,王刘庆,李祖梁,王 多,王 蒙,*
(1.北京农业质量标准与检测技术研究中心,农业农村部农产品质量安全风险评估实验室(北京),农产品产地环境监测北京市重点实验室,北京 100097;2.新疆农业大学食品科学与药学学院,新疆 乌鲁木齐 830052)
交链孢菌(Alternariaspp.)是污染农产品及其制品最普遍的真菌之一,广泛分布于自然界,其中互隔交链孢(Alternaria alternata)是主要的病原菌,其不仅能够侵染多种农作物,引起植物病害(如番茄黑斑病等),而且能够产生交链孢毒素。检出率较高的交链孢毒素包括细交链孢酮酸(tenuazonic acid,TeA)、交链孢酚(alternariol,AOH)、交链孢酚单甲醚(alternariol monomethyl ether,AME)等[1-3]。交链孢毒素对动植物具有一定的潜在危害,其中AOH具有遗传毒性和致突变性,还可促使机体产生氧化应激反应[4-6]。TeA与AOH、AME具有协同作用,且均具有动物急性毒性[7]。欧洲食品安全局的数据显示随膳食摄入的AOH对公众健康存在风险[8-10],研究显示河南省林县地区食管癌高发可能与粮食作物中交链孢毒素污染,特别是AOH及其甲基化产物AME含量较高有关[11-12]。目前食品中的交链孢毒素污染已成为一个重要的公共卫生问题,因此掌握有效抑制交链孢菌生长及其产毒的防控方法具有重要意义。
臭氧作为强氧化剂,可迅速破坏有机物中的双键,形成含双键少、分子质量较低的物质;且低剂量臭氧作用于粮食、果蔬等农产品表面即可起到杀菌消毒的作用[13]。臭氧可自动分解为氧气,无毒性残留,早在1997年,美国食品药品监督管理局就将臭氧作为一种普遍认为安全的方法应用于食品加工过程中,目前臭氧已在食品加工贮藏、果蔬保鲜、废水处理等领域得到广泛应用[14]。臭氧处理还可以杀灭真菌、有效破坏毒素的化学结构,从而实现降解毒素的目的。近年来,臭氧用于防控产毒真菌、降解真菌毒素的研究多有报道。Mylona等研究发现臭氧气体不仅在体外可抑制轮枝镰刀菌菌丝的生长和孢子的萌发,同时可抑制在玉米粉中伏马毒素(fumonisin B,FB)的积累[15]。Savi等的研究表明臭氧处理可以抑制禾谷镰刀菌的生长,同时可有效降解小麦粒中的脱氧雪腐镰刀菌烯醇(deoxynivalenol,DON)[16]。此外,还有研究报道臭氧用于降解花生及红辣椒中的黄曲霉毒素B1(aflatoxin B1,AFB1)[17-18]、苹果汁中的展青霉素[19]、玉米中的玉米赤霉烯酮(zearalenone,ZEN)以及赭曲霉毒素(ochratoxin A,OTA)[20]等。一些臭氧对互隔交链孢影响的研究也表明臭氧处理可抑制互隔交链孢的孢子萌发及菌丝生物量生长[21-22]。然而,鲜见臭氧处理拮抗交链孢产毒以及其直接降解毒素的研究。鉴于此,本研究不仅考察不同剂量的臭氧处理对体外互隔交链孢生长及产毒能力的影响;而且通过接种番茄研究臭氧处理后互隔交链孢在番茄活体内的致病能力及产毒情况,同时分析了臭氧对交链孢毒素的直接降解效果,以期为利用臭氧防控互隔交链孢侵染及其毒素积累提供参考。
1 材料与方法
1.1 材料与试剂
红熟番茄(Solanum lycopersicumcv. Shengfan15)采于北京天安农业有限公司的番茄园,当天运回实验室,选择大小均一、同一成熟度、果形端正、无机械损伤和病虫害的番茄果实作为实验用果。将番茄果实经体积分数1%次氯酸钠溶液浸泡1 min,随后冲洗,自然晾干备用。
交链孢毒素标准品(AOH、AME、TeA)(纯度均大于99%) 美国Sigma-Aldrich公司;乙腈、甲醇、甲酸、乙酸铵(均为质谱级) 美国Fisher公司;马铃薯葡萄糖琼脂(potato dextrose agar,PDA) 美国Becton公司。
1.2 仪器与设备
AcquityTM超高效液相色谱串联质谱(ultra performance liquid chromatography-tandem mass spectrometry,UPLC-MS/MS)仪、电喷雾电离(electron spray ionization,ESI)检测器 美国Waters公司;3-30k台式高速冷冻离心机 德国Sigma公司;N-EVA-112P氮吹仪 美国Organomation公司;A11分析研磨机、漩涡混合器 德国IKA公司;Milli-Q A10超纯水器 美国Millipore公司;QJ-8006K臭氧发生器 广州铨聚臭氧科技有限公司;APES-O3臭氧检测仪 深圳安帕尔科技有限公司;S-3400N扫描电子显微镜 日本Hitachi公司。
1.3 方法
1.3.1 互隔交链孢培养及孢子悬浮液配制
本研究所用的互隔交链孢为本实验室前期从番茄上分离纯化得到的菌株,将互隔交链孢接种于PDA培养基上,25 ℃条件下恒温避光培养10 d。然后用体积分数为0.05%的Tween-80溶液冲洗平板,采用血球计数板法制成浓度约为1×105个/mL的孢子悬浮液。
1.3.2 样品制备
1.3.2.1 臭氧处理互隔交链孢孢子悬浮液
参考文献[23]的方法并有所修改。分别吸取2 mL交链孢菌孢子悬浮液置于25 mL小烧杯中(烧杯直径为5 cm),将装有孢子悬浮液的小烧杯放入已消毒的密闭贮藏箱(48 cm×28 cm×28 cm)中,同时将臭氧检测器置于贮藏箱中,在室温条件下通入臭氧气体,控制臭氧发生器使箱内臭氧质量浓度分别为5、10、20 mg/L,处理30 min。处理结束后回收烧杯内的孢子悬浮液,以不经臭氧处理的孢子悬浮液作为对照,分别接种于PDA培养基及番茄果实上。
接种PDA培养基:分别吸取对照及不同质量浓度臭氧处理后的交链孢菌孢子悬浮液20 μL接种于PDA平板上,25 ℃条件下恒温避光培养7 d,每组接种5 个平板,分别于第3、5、7天用游标卡尺十字交叉测量菌落直径,并于第7天取1/4培养基进行交链孢毒素的提取及测定,实验重复3 次。
接种番茄果实:接种前用直径3 mm、已灭菌的不锈钢钉子在番茄赤道部位刺入1 个伤口(3 mm×3 mm),待伤口表面晾干后分别吸取对照及不同质量浓度臭氧处理后的交链孢菌孢子悬浮液20 μL接种于番茄果实上,每组设置15 个番茄,接种后放置在室温为20 ℃的环境中贮藏7 d,分别于第3、5、7天用游标卡尺十字交叉测量病斑部位的直径,并于第7天在病斑部位处取样。将病斑部位样品用研磨机混合匀浆,贮存于-20 ℃待测定交链孢毒素含量,实验重复3 次。
1.3.2.2 臭氧处理交链孢毒素标准品
参考文献[14]的方法并有所修改。将TeA、AOH、AME 3 种交链孢毒素标准品用乙腈稀释定容,配制成质量浓度为1 μg/mL的标准溶液,分别吸取1 mL标准溶液置于25 mL小烧杯中(烧杯直径为5 cm)。将装有毒素标准溶液的小烧杯放入已消毒的密闭贮藏箱(48 cm×28 cm×28 cm)中,同时将臭氧检测器置于贮藏箱中,在室温条件下通入臭氧气体,控制臭氧发生器使箱内臭氧质量浓度分别为2、5、10、20、40 mg/L,处理30 min;另外,固定臭氧质量浓度为20 mg/L,分别处理毒素标准溶液15、30、45、60、90、120 min。处理结束后,回收烧杯内的毒素标准溶液,利用UPLC-MS/MS进行测定,以不经臭氧处理的毒素标准溶液作为对照,每个处理组设置3 个平行,实验重复3 次,考察不同臭氧质量浓度及处理时间对交链孢毒素的降解作用。臭氧处理交链孢毒素降解率按下式计算。

式中:c(A,t)为交链孢毒素经臭氧处理时间t后的含量/(μg/kg);c(A,0)为交链孢毒素的初始含量/(μg/kg)。
1.3.3 扫描电子显微镜观察互隔交链孢的微观形态
参考文献[24]的方法并有所修改。为进一步研究臭氧对互隔交链孢孢子和菌丝形态的影响,对其进行扫描电子显微镜观察。选择10、20 mg/L臭氧处理后的孢子悬浮液接种于PDA培养基上,25 ℃条件下恒温避光培养24 h后在菌落周围插入灭菌的玻璃片,培养24 h后取出玻璃片。将长有交链孢菌的玻璃片置于戊二醛溶液中固定2~12 h,用0.1 mol/L pH值为6.8的磷酸盐缓冲液冲洗2 次,每次10 min;依次用体积分数20%、50%、70%、80%、90%乙醇溶液梯度洗脱,每次15 min,再用无水乙醇洗脱2 次,每次15 min;最后用叔丁醇置换,每次30 min,共置换3 次,最后一次置于4 ℃冰箱里,待叔丁醇凝固后冷冻干燥,随后进行扫描电子显微镜观察,以不经臭氧处理组为对照,实验重复3 次。
1.3.4 交链孢毒素含量的测定
交链孢毒素的提取及含量测定参考本课题组前期研究的方法[25],并略有修改。取番茄病斑样品(5 g)及1/4 PDA培养基样品于50 mL离心管内,加入20 mL含有体积分数1%甲酸的乙腈溶液,漩涡混匀,经150 r/min常温振荡提取30 min,漩涡振荡后经高速冷冻离心机离心(1 000 r/min、10 min、10 ℃),离心完成后取1 mL上清液于40~50 ℃条件下氮气吹至近干,随后用1 mL乙腈溶液(体积比3∶7)复溶,混匀后过0.22 μm PTFE微孔滤膜,经UPLC-MS/MS测定,每个样品设定3 个平行,实验重复3 次。
1.4 数据统计及分析
实验中所有数据的方差分析及多重比较均采用SPSS 23.0软件进行,用Duncan’s法进行差异显著性检验(P<0.05),采用Gaphpad prism软件作图。
2 结果与分析
2.1 臭氧处理对互隔交链孢菌落直径的抑制
表1所示为臭氧处理互隔交链孢孢子悬浮液后接种到PDA培养基上,随着贮藏时间的延长不同臭氧质量浓度处理对互隔交链孢菌落直径的抑制情况。从表1可以看出,臭氧气体对互隔交链孢菌丝的生长有较大程度的延缓和抑制作用,处理组的菌落比对照组生长显著减慢(P<0.05)。同时可以看出,抑制作用与臭氧质量浓度有关,随着臭氧质量浓度的增加,其对互隔交链孢菌落直径的抑制作用明显加强,互隔交链孢菌丝生物量积累逐渐减小。在培养第7天,对照组的菌落直径为6.88 cm,此时臭氧质量浓度为10 mg/L的处理组菌落直径为5.58 cm,臭氧质量浓度为20 mg/L的处理组菌落直径仅为5.10 cm,显著低于对照组及低质量浓度的臭氧处理组(P<0.05),其对互隔交链孢生长的抑制率分别为18.9%和25.9%。
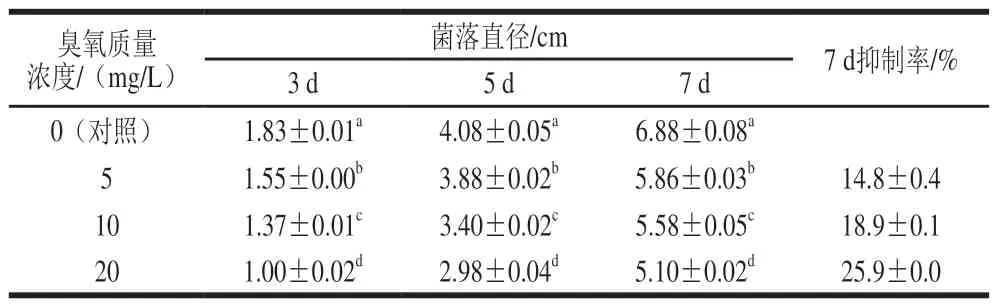
表 1 臭氧处理对交链孢菌生长的抑制情况Table 1 Inhibitory effect of ozone on mycelial growth of A. alternata
臭氧直接作用于互隔交链孢孢子悬浮液,导致其生长受到抑制,这可能是因为臭氧的强氧化作用直接使分生孢子失活,使其完全丧失萌发能力;或者臭氧破坏了分生孢子的细胞结构,从而使健康的分生孢子数量减小,进而导致菌落生长缓慢。本研究结果与文献报道一致,Ames等[21]将互隔交链孢长期置于低含量臭氧气体(150 nL/L)中,随着暴露时间的延长,互隔交链孢的菌落直径生长逐渐减慢、分生孢子数量逐渐减小,在100 d时分生孢子全部失活。Tzortzakis等也研究了不同浓度的臭氧熏蒸对交链孢菌的菌丝生物量及分生孢子的影响,结果表明臭氧处理后互隔交链孢的菌丝生物量及分生孢子活力均受到显著抑制,且随着臭氧质量浓度的增加抑制作用显著增强;同时,他们指出臭氧对真菌孢子的抑制作用也与孢子形态、水分含量、生长基质有关,具体的抑制机理还有待研究[22]。
2.2 臭氧处理对互隔交链孢菌丝及孢子形态的影响
为了进一步考察臭氧处理对互隔交链孢造成的破坏,采用扫描电子显微镜观察臭氧处理后的互隔交链孢菌丝及孢子的形态变化情况。如图1所示,与对照相比,臭氧处理后的互隔交链孢形态发生异常。正常的互隔交链孢孢子呈完整、丰满的卵形、倒梨形或近似椭球形,表面光滑或具微刺,菌丝表面浓厚光滑,自然伸长。经10 mg/L臭氧处理30 min后互隔交链孢孢子发生很明显的凹陷、褶皱,菌丝变得干瘪、萎缩、粗糙,同时出现断裂的情况;臭氧质量浓度增大到20 mg/L时,菌丝和孢子形态异常程度也逐渐增大,除了明显的凹陷和褶皱外,孢子表面还发生破裂,菌丝更加干瘪粗糙,且出现不规则的折叠、皱缩,同时有些孢子和菌丝表面出现小囊泡,由此可见臭氧对互隔交链孢的菌丝及孢子的破坏作用与臭氧质量浓度有关。
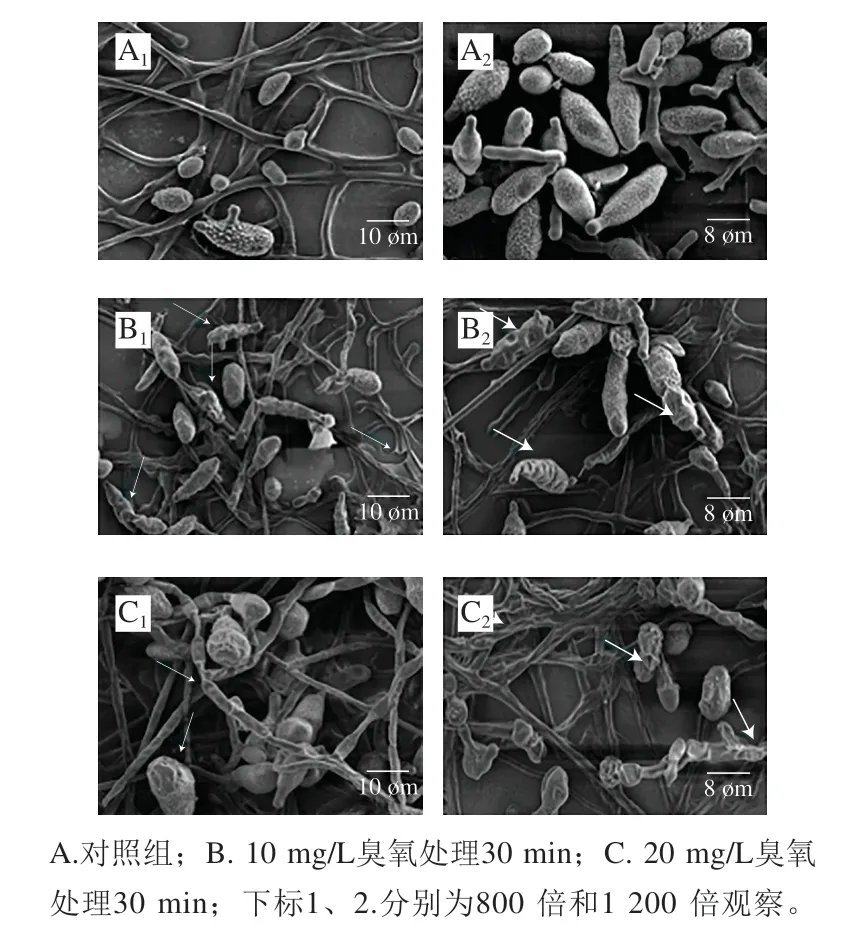
图 1 扫描电子显微镜观察臭氧处理后互隔交链孢的微观形态Fig. 1 SEM observation of A. alternata after ozone treatment
当真菌暴露在不利于生长的环境(如化学处理、低温、高湿等)中就会出现分生孢子减少、菌落生长缓慢等现象[26-27]。虽然目前尚鲜见臭氧对互隔交链孢形态学影响的研究报道,但Savi等[28]研究了臭氧对镰刀菌(Fusarium)、曲霉菌(Aspergillus)以及青霉菌(Penicillium)等真菌形态学的影响,以上3 种真菌经过臭氧处理后在形态上均表现出异常,菌丝出现变形、破裂。郭宇欢等[23]将灰葡萄孢(Botrytis cinerea)暴露于臭氧气体中,发现菌丝表面出现沟壑,说明臭氧处理对灰葡萄孢菌丝造成伤害。在本研究中,一方面,臭氧对互隔交链孢的破坏可能是因为强氧化作用直接导致大量孢子及菌丝发生破坏,进而引起真菌死亡;另一方面,臭氧的强氧化作用可能使细胞壁上的蛋白质、几丁质、葡聚糖等物质发生化学作用,改变细胞壁的完整性,使细胞内容物释放,进而导致真菌细胞遭到破坏[29]。
2.3 臭氧处理对体外互隔交链孢产毒能力的影响
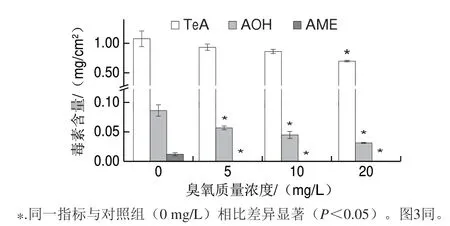
图 2 经臭氧处理后互隔交链孢在体外PDA培养基上所产毒素含量Fig. 2 Concentrations of various mycotoxins produced by A. alternata with ozone treatment when cultured on PDA plates
不同质量浓度臭氧处理互隔交链孢孢子悬浮液后接种于PDA培养基上,25 ℃条件下避光生长7 d,第7天进行交链孢毒素的提取及含量测定,图2所示为体外PDA培养基上交链孢毒素含量的变化情况。对照组互隔交链孢高产TeA(1.08 mg/cm2),AOH(0.071 mg/cm2)、AME(0.009 mg/cm2)含量相对较低。本研究结果表明臭氧处理可显著抑制A.alternata产TeA、AOH、AME这3 种交链孢毒素,且与臭氧质量浓度呈正相关。在低质量浓度臭氧(5 mg/L)处理时,TeA含量为0.88 mg/cm2,AOH含量为0.065 mg/cm2,均低于对照组,而AME不再产生。随着臭氧质量浓度的增加,TeA和AOH的含量也逐渐降低,当臭氧质量浓度增加至20 mg/L时,TeA含量减少至0.71 mg/cm2,与对照组相比减少了34.3%;AOH的含量减少至0.033 mg/cm2,比对照减少了53.5%。
交链孢毒素含量的降低可能是因为互隔交链孢经臭氧处理后菌丝和孢子受到损伤,导致总的生物量减少,进而导致产生毒素的含量有所降低。目前尚鲜见臭氧处理对互隔交链孢体外产毒影响的研究,但有臭氧对其他真菌产毒的文献报道。Mylona等[15]将长有轮枝镰刀菌的培养基暴露于臭氧气体中(100×10-6、200×10-6,1 h),10 d后,与对照相比轮枝镰刀菌FB产量显著降低。关于臭氧能抑制真菌产毒的机理,有研究认为是因为臭氧作用使真菌的细胞膜完整性以及参与多种化学反应的酶遭到破坏[30],而本研究也发现臭氧能够有效地破坏互隔交链孢细胞结构,进而减少毒素的合成,然而关于臭氧抑制互隔交链孢产毒的详细机制还有待进一步研究。
2.4 臭氧处理对互隔交链孢致病能力及产毒能力的影响
互隔交链孢是引起番茄黑斑病的主要病害之一,并且会在番茄果实体内积累产生交链孢毒素,这对番茄及其加工制品的食用安全造成较大威胁。基于此,本研究选择番茄为基质,将臭氧处理后的互隔交链孢接种到健康番茄体内,观察互隔交链孢在番茄上生长的病斑直径大小以及产毒的变化情况。
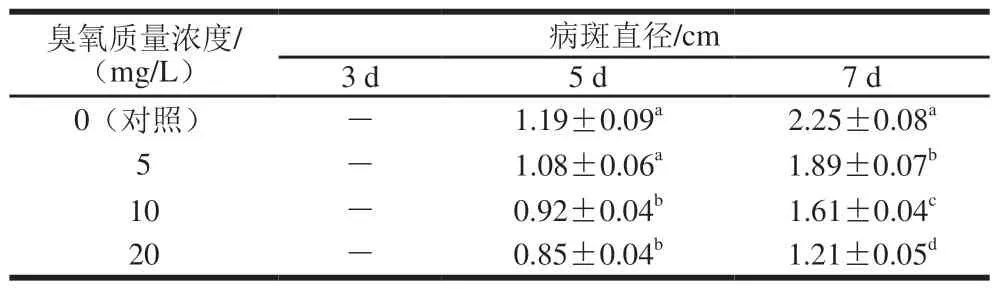
表 2 经臭氧处理后的交链孢菌在番茄上的生长情况Table 2 Mycelial growth of A. alternata with ozone treatment on tomato fruit
不同质量浓度臭氧处理互隔交链孢后接种到番茄果实上,第3、5、7天的病斑直径如表2所示。随着生长时间的延长,互隔交链孢病斑直径逐渐扩大,臭氧处理导致病斑延伸较对照显著放缓(P<0.05),且不同质量浓度的臭氧处理组之间也表现出显著差异(P<0.05)。随着臭氧质量浓度的增大,互隔交链孢在番茄果实上生长受抑制程度逐渐增大,说明臭氧处理对互隔交链孢致病力有较明显的抑制作用,且臭氧质量浓度与致病力呈负相关。龙君[31]用不同质量浓度(50、100、150、200 mg/m3)臭氧气体熏蒸意大利青霉(Penicillium italicum),随后接种到柑橘上观察柑橘的发病情况,实验结果表明臭氧熏蒸后柑橘青霉菌分生孢子的致病力与对照组相比有显著差异,臭氧气体既能够抑制柑橘发病,也能够延缓病斑的扩展。这与本研究结果类似,臭氧处理组均表现出对果蔬病原菌的拮抗作用,能够抑制病害的发生。
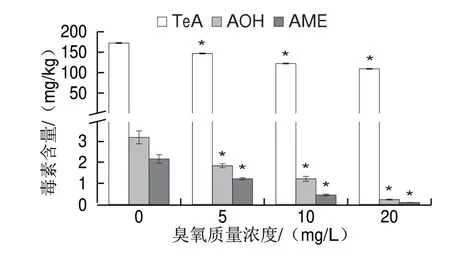
图 3 臭氧处理后番茄病斑部位的交链孢毒素含量Fig. 3 Concentrations of various Alternaria toxins in lesions of tomato fruit inoculated with A. alternata with ozone treatment
图3所示为臭氧处理后的互隔交链孢在番茄果实上生长第7天的产毒情况。对照组互隔交链孢在番茄基质上产TeA、AOH、AME的含量分别为172.50、3.18、2.22 mg/kg。与对照组相比,臭氧处理组中3 种交链孢毒素的含量均有所下降,且随着臭氧质量浓度的增大,毒素含量显著降低(P<0.05),说明臭氧处理对互隔交链孢产毒能力具有干预作用,且随着臭氧质量浓度的增大,抑制产毒能力逐渐增强。然而,臭氧处理对互隔交链孢产3 种毒素的抑制能力有所差异,经20 mg/L臭氧处理的番茄果实贮藏7 d后,TeA的含量为110.2 mg/kg,比对照组减少了36.1%;而AOH和AME的含量分别为0.32 mg/kg和0.15 mg/kg,分别比对照组减少了89.9%和93.2%,可见臭氧处理抑制该菌株产AOH和AME毒素的作用更明显。这说明臭氧处理对互隔交链孢的菌丝和孢子造成破坏,导致其不能在番茄上正常生长,进而引起交链孢毒素的积累减少;而不同交链孢毒素之间抑制率的明显差异又显示出臭氧处理对互隔交链孢产毒的影响不单是缘于对互隔交链孢的抑制,也可能存在其他机制影响毒素的合成。
番茄果实富含酚酸,主要有绿原酸、咖啡酸、对香豆酸、阿魏酸、4-羟基苯甲酸等[32],同时有研究表明互隔交链孢接种到番茄上会引起酚酸含量的变化,而且体外抑菌实验也表明绿原酸及咖啡酸对互隔交链孢生长的抑制率分别是16%、30%[33]。Pane等的研究表明花椒中多酚提取物对采后互隔交链孢侵染樱桃番茄具有抑制作用[34]。酚酸不仅抑制病原真菌生长,对真菌毒素的积累也具有抑制作用,本课题组前期的研究结果表明,对香豆酸、咖啡酸可以显著抑制互隔交链孢的生长及TeA的产生[35]。另外也有研究表明0.5 mmol/L绿原酸及阿魏酸可使轮枝镰刀菌产T-2毒素分别减少23.1%及26.5%[36]。酚酸是植物防御体系的组成部分,互隔交链孢在番茄上的致病力及产毒能力受到抑制可能是由于臭氧和酚酸协同作用的结果,但是不同酚酸成分对交链孢毒素的抑制效果及防御机理还需进一步研究。
2.5 臭氧处理对交链孢毒素的降解作用

图 4 臭氧处理交链孢毒素的降解效果Fig. 4 Degradation of Alternaria toxins by ozone treatment
臭氧因其强氧化作用可以降解真菌毒素或改变其化学结构,使其毒性基团遭到破坏,从而起到解毒作用。目前众多研究都表明臭氧对谷物中的AFB1、OTA、ZEN等具有良好的降解效果,但是对交链孢毒素的降解效果鲜见报道。图4为不同臭氧质量浓度及处理时间对TeA、AOH、AME 3 种交链孢毒素的降解效果。从图4A可以看出,TeA、AOH、AME 3 种交链孢毒素的降解率随臭氧质量浓度的增大而逐渐升高,其中TeA经20 mg/L臭氧处理30 min后已检测不到;AOH在较低臭氧质量浓度下降解效果就比较显著,臭氧质量浓度为2 mg/L时降解率为56.0%,臭氧质量浓度增大至40 mg/L时降解率为75.0%,降解率变化幅度不明显;而AME在较低臭氧质量浓度时降解效果不明显,臭氧质量浓度增大至40 mg/L时降解率为63.3%。从图4B中可以看出,当臭氧质量浓度固定为20 mg/L时,随着处理时间的延长,交链孢毒素的降解率也逐渐增大,对TeA的降解效果最显著,在15 min时降解率达63.5%,随着时间延长,到30 min时可被完全降解;AOH在45 min时降解率为78.8%,在60 min时降解率为79.5%,在此时间段降解率变化不显著,随着时间进一步延长,至120 min时降解率达91.9%;AME在较短时间内降解效果较差,45 min时降解率仅为47.6%,长时间臭氧处理可较大程度上降解AME,臭氧处理120 min时其降解率达到90.0%。
从两组实验结果来看,处理时间对3 种交链孢毒素降解效率的影响占主导因素,除了TeA在较短时间就可降解以外,AOH及AME在20 mg/L处理60 min时降解率即可达70%以上。与文献报道的臭氧处理其他真菌毒素相比,3 种交链孢毒素的臭氧处理时间较长、所需臭氧质量浓度较高。Qi Lijun等[20]研究了不同质量浓度臭氧及时间处理ZEN及OTA的效果,5 mg/L臭氧处理ZEN标准溶液5 s,其降解率超过90%;10 mg/L臭氧处理OTA标准溶液10 s,其降解率接近50%。王莉等[14]用质量浓度为10 mg/L的臭氧气体降解DON,结果表明随着处理时间的延长,DON的降解率也随之提高,处理5 min后DON降解率达到88%。由此可见,臭氧质量浓度、处理时间、毒素的初始浓度、水分含量、pH值等因素都会不同程度地影响臭氧降解真菌毒素的效果[37]。另外,臭氧对真菌毒素的降解效果也与其结构和特性有关,臭氧具有强氧化作用,一般会攻击双键结构发生加成反应,也可能会破坏苯环结构。现有研究认为关于臭氧降解AFB1、DON、ZEN的机理主要为Criegee机制[14,38-39],但是关于交链孢毒素的臭氧降解产物鉴定、降解路径解析、产物安全性评价等方面还需开展深入研究。
3 结 论
臭氧作为一种强效的氧化剂,因其极易分解生成氧气、无污染残留等优点,已广泛应用于食品加工贮藏、果蔬保鲜等领域。本研究结果表明臭氧处理可导致互隔交链孢菌丝干瘪萎缩和孢子凹陷褶皱,从而导致其在离体时表现为生长受到抑制,番茄果实上表现为致病力减弱,进而引起互隔交链孢产TeA、AOH、AME 3 种毒素能力的降低。同时,臭氧处理还能降解3 种交链孢毒素,随臭氧质量浓度的增加及作用时间的延长,其对交链孢毒素的降解能力逐渐增加,如TeA经过20 mg/L臭氧处理15 min就可以被大量降解,降解率为63.5%,AOH及AME在20 mg/L时臭氧处理60 min,降解率可达70%以上。此外,众多研究已表明臭氧对病原真菌的生长及其毒素积累均具有较强的抑制作用。因此,臭氧技术可以作为农产品及其制品中互隔交链孢及其毒素污染的防治手段。本研究初步探索了臭氧技术对互隔交链孢及其产毒能力的影响,下一步将深入研究臭氧抑制互隔交链孢产毒的机理,鉴定臭氧降解交链孢毒素的降解产物,解析降解路径并评价降解产物的安全性,以期为开展果蔬贮藏加工过程中利用臭氧防控交链孢毒素污染提供科学依据和技术支撑。
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
煤气与热力(2021年10期)2021-12-02
浙江农业学报(2021年5期)2021-05-31
新农业(2017年15期)2017-09-11
中学生数理化·八年级物理人教版(2017年12期)2017-04-18
上海农业学报(2017年4期)2017-04-10
中国蔬菜(2016年8期)2017-01-15
中国感染控制杂志(2015年7期)2015-12-13
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年1期)2015-07-07
