
高温及其联合亚硒酸钠调控西兰花芽苗生理及异硫氰酸酯代谢研究
2019-11-28 06:57程雨薇沈敏熙朱振宇方维明尹永祺
食品工业科技 2019年22期
韩 宇,程雨薇,沈敏熙,朱振宇,程 超,方维明,尹永祺
(扬州大学食品科学与工程学院,江苏扬州 225127)
近年来,富含异硫氰酸酯(Isothiocyanates,ITCs)的功能性食品日益受到关注。ITCs是蔬菜中硫代葡萄糖苷(Glucosinolates,简称硫苷)在黑芥子酶(Myrosinase,MYR,EC 3.2.1.147)作用下形成的水解产物[1],对人体具有抗氧化和抗癌等多种生理功效[2]。ITCs已被证实是抗癌作用最强的天然植物活性成分,其可通过抑制阶段I酶和激活阶段II酶来发挥其抗癌功能[3-4]。流行病学研究显示,食用西兰花等芸薹属蔬菜可预防胰腺癌、肺癌和直肠癌等多种癌症的发生,这主要与硫苷的水解产物ITCs有关[5]。
植物籽粒发芽尤其是逆境胁迫条件下发芽后其硫苷含量显著增加,内源黑芥子酶被激活,因此,通过控制发芽条件以富集芽苗中ITCs已成为一种行之有效的方法。研究发现,植物发芽期间经盐胁迫[6]、硫酸锌胁迫[7]、低氧胁迫[8]以及脱落酸处理[9],其芽苗中ITCs含量显著提高。硒和硫为同族元素,其在原子大小、化学性质方面相似,新陈代谢中硒可以替代硫起作用,以非特异性的方式取代硫。研究显示亚硒酸钠(Na2SeO3)处理可以提高芽苗中硫苷的含量[10-12]。同时,甘蓝根经盐胁迫及其联合Na2SeO3处理均可促进ITCs富集[13]。然而,高温胁迫尤其是高温联合Na2SeO3处理对于芽苗生理及ITCs代谢的影响未见报道。由于硫苷是防癌物质的前体,与硒交换硫可能进一步提高硫代葡萄糖苷水解产物的生物活性,因此研究硒对硫苷的富集具有重要意义。
本研究以西兰花为试材,研究高温及其联合Na2SeO3处理下芽苗主要生理生化和ITCs代谢的变化,以期初步揭示高温及其联合Na2SeO3处理调控ITCs代谢的作用机制,为开发富含ITCs芽类营养食品提供技术支撑和理论依据,推动芽苗菜的工业化生产。
1 材料与方法
1.1 材料与仪器
西兰花种子(BrassicaoleraceaL.,优秀) 购于2018年,产自日本坂田公司,封装于密闭容器中,4 ℃保存备用;亚硒酸钠、烯丙基硫苷(纯度>95%)、萝卜硫素(纯度>99%)生化试剂 均为美国Sigma公司;1,2-苯二硫醇(纯度>99%) 生化试剂,东京化成工业株式会社;其余试剂均为国产分析纯;过氧化氢试剂盒A064、总抗氧化能力检测试剂盒A015-1、葡萄糖测定试剂盒 为南京建成生物工程研究所。
pGX-150型智能光照培养箱 宁波海曙赛福实验仪器厂;UV-7504C型紫外可见分光光度计 上海欣茂仪器有限公司;DK-S12型电热恒温水浴锅 上海森信实验仪器有限公司;DICO型台式离心机 Thermo Fisher有限公司;XW-80C型漩涡混合器 上海医科大学仪器厂。
1.2 实验方法
1.2.1 亚硒酸钠浓度筛选 称取西兰花种子12 g,置于100 mL 1%(v/v)NaClO水溶液中20 ℃浸泡消毒15 min,消毒种子用蒸馏水冲洗至pH中性后于蒸馏水中30 ℃浸泡4 h,其后置于铺有蛭石的透明盒中,于30 ℃培养箱内发芽,16 h光照/8 h黑暗,分别用20 mL的0、10、50、100、200 μmol/L的Na2SeO3溶液喷施,发芽4 d后,测定各处理组西兰花芽苗中异硫氰酸酯含量。
1.2.2 高温及其联合Na2SeO3处理试验 按1.2.1中发芽方法,分别设以下处理:对照处理(蒸馏水)、高温处理(60 ℃热激20 min)、高温联合Na2SeO3处理(60 ℃热激20 min,0.1 mmol/L Na2SeO3)。芽苗共发芽6 d,第1 d均喷施20 mL去离子水催芽,之后每隔24 h将热激组芽苗置于60 ℃恒温箱热激处理20 min[14-15]并对不同处理组分别喷施20 mL不同处理液至培养结束,发芽期间4、6 d取样测定相关指标。
1.2.3 指标测定与方法
1.2.3.1 形态特征 从各处理组中随机选取10株西兰花芽苗,摆放整齐后,观察其形态特征。
1.2.3.2 芽长及鲜重 随机选取30株西兰花芽苗,用游标卡尺测定其芽长,并用精密电子天平进行鲜重的测定,计算单株芽苗鲜重。
1.2.3.3 过氧化氢、丙二醛含量及总抗氧化能力的测定 过氧化氢含量利用过氧化氢试剂盒测定;参照硫代巴比妥酸法[16]测定丙二醛含量;总抗氧化能力(T-AOC)利用试剂盒测定。
1.2.3.4 ITCs、硫苷含量测定 参照Jiao等[17]的方法并修改。准确称取0.2 g芽苗,加入3 mL蒸馏水研磨,匀浆液于40 ℃水浴3 h后加入3 mL二氯甲烷振荡萃取30 min,10000×g离心15 min,取下清液100 μL依次加入2 mL甲醇、1.8 mL 50 mmol/L硼酸缓冲液和0.2 mL 7 mmol/L 1,2-苯二硫醇的甲醇溶液,混合后置于65 ℃水浴1 h后于365 nm处测定吸光值。用萝卜硫素作标准曲线,计算样品ITCs含量。硫苷含量参照Wei等[18]的方法测定。
1.2.3.5 黑介子酶活性 参照Burow等[19]的方法。取0.2 g芽苗,用3 mL 0.1 mol/L pH6.5磷酸盐缓冲液冰浴研磨,4 ℃下10000×g离心15 min。取上清液0.5 mL与0.5 mL 0.25 mmol/L烯丙基硫苷溶液混合,于37 ℃水浴反应15 min后沸水浴5 min以灭酶,用葡萄糖试剂盒测定葡萄糖含量。以每分钟被黑芥子酶转化生成1 nmol葡萄糖为1个酶活力单位(U/mg protein)。
1.3 数据处理
试验结果以平均值±标准差表示。数据采用统计分析软件SPSS 18.0进行统计分析,均值间比较采用Tukey多重比较,在0.05水平上进行显著检验(P<0.05)。
2 结果与分析
2.1 亚硒酸钠浓度筛选
如图1所示亚硒酸钠浓度为100 μmol/L时西兰花芽苗中异硫氰酸酯含量显著高于其他浓度处理(P<0.05),故本实验选择0.1 mmol/L亚硒酸钠溶液作为处理液。

图1 不同浓度亚硒酸钠对西兰花芽苗中异硫氰酸酯含量的影响Fig.1 Effects of different concentrations of Na2SeO3on the content of isothiocyanate in broccoli sprouts
2.2 高温及其联合Na2SeO3处理对西兰花芽苗生理代谢的影响
2.2.1 形态特征 图2直观显示西兰花芽苗经高温胁迫及其联合Na2SeO3处理后,其生长状态均受到影响,且经高温胁迫联合Na2SeO3处理的西兰花芽苗受胁迫情况更严重,相较于对照,其芽苗及叶片的大小均明显受到抑制。

图2 高温及其联合Na2SeO3处理对西兰花芽苗形态表征的影响Fig.2 Effects of high temperature and its combinedwith Na2SeO3 on morphology of broccoli sprouts注:W:蒸馏水(空白对照);H:高温;H-Se:高温+Na2SeO3,图3~图9同。
2.2.2 芽长及鲜重 由图3可看出,发芽期间各处理下的西兰花芽苗芽长及鲜重均随发芽时间延长呈增长趋势,发芽4 d时,各处理间芽长均具有显著差异(P<0.05,图3A),热激处理下鲜重显著低于对照处理(P<0.05,图3B)。发芽6 d时,高温胁迫及其联合Na2SeO3处理的芽苗芽长均显著低于对照处理(P<0.05)。高温胁迫联合Na2SeO3处理发芽4、6 d时芽长分别为对照的79%和57%;发芽6 d时,高温胁迫联合Na2SeO3处理的芽苗芽长及鲜重与高温胁迫处理的无显著差异(P>0.05),但高温胁迫联合Na2SeO3处理的芽苗鲜重显著低于对照处理(P<0.05),表明高温及其联合Na2SeO3处理均显著抑制了西兰花芽苗的生长。

图3 高温及其联合Na2SeO3处理对西兰花芽苗芽长(A)及鲜重(B)的影响Fig.3 Effects of high temperature and its combined treatmentwith Na2SeO3 on the sprout length(A)and fresh weight(B)of broccoli sprouts注:不同字母表示不同处理间芽苗的芽长及鲜重显著差异,P<0.05;图4~图9同。
2.2.3 过氧化氢含量 如图4,发芽期间各处理下西兰花芽苗中过氧化氢含量均随发芽时间延长呈下降趋势。发芽4 d时,相较对照处理,高温及其联合Na2SeO3处理的芽苗中过氧化氢含量均无显著差异(P>0.05);发芽6 d时,高温胁迫联合Na2SeO3处理的芽苗中过氧化氢含量较空白对照显著提高(P<0.05),其含量较空白对照提高了57%,而高温胁迫处理与空白对照处理无显著差异(P>0.05)。
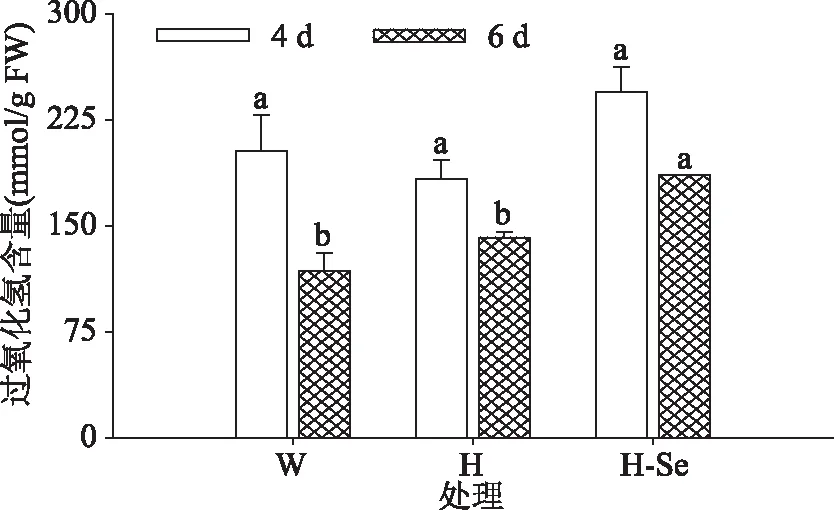
图4 高温及其联合Na2SeO3处理对西兰花芽苗过氧化氢含量的影响Fig.4 Effects of high temperature and its combined treatmentwith Na2SeO3 on hydrogen peroxide content in broccoli sprouts
2.2.4 丙二醛含量 植物器官衰老或在逆境下遭受伤害,往往发生膜脂过氧化作用,丙二醛(MDA)是膜脂过氧化的最终分解产物,其含量可以反映植物遭受逆境伤害的程度,同时,它的产生还能加剧膜的损伤,间接地反映了植物组织的抗氧化能力的强弱[16]。如图5,发芽期间各处理下西兰花芽苗中丙二醛含量均随发芽时间延长呈下降趋势。发芽4 d,较对照而言,高温胁迫及其联合Na2SeO3处理的芽苗中丙二醛含量均显著增加(P<0.05),发芽6 d时,高温胁迫与高温联合Na2SeO3处理的芽苗中丙二醛含量无显著差异(P>0.05),但仍均显著高于空白对照(P<0.05),表明芽苗经高温及其联合Na2SeO3处理后其生长受到了抑制。
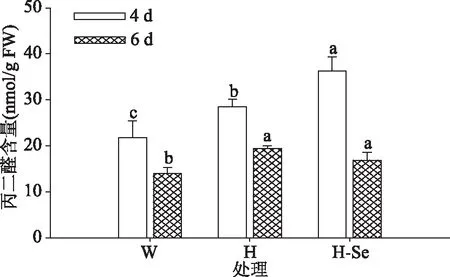
图5 高温及其联合Na2SeO3处理对西兰花芽苗丙二醛含量的影响Fig.5 Effects of high temperature and its combined treatmentwith Na2SeO3 on malondialdehyde content in broccoli sprouts
2.2.5 总抗氧化能力 如图6,发芽期间各处理下西兰花芽苗中总抗氧化能力均随发芽时间延长呈下降趋势。发芽4、6 d时,高温胁迫及其联合Na2SeO3处理的芽苗中总抗氧化能力均较空白对照显著提高(P<0.05),4 d时其总抗氧化能力较空白对照分别提高了29%和57%,而6 d时则较空白对照分别提高了25%和45%;发芽4、6 d时,高温胁迫及其联合Na2SeO3处理的芽苗中总抗氧化能力无显著差异(P>0.05)。

图6 高温及其联合Na2SeO3处理对西兰花芽苗总抗氧化能力的影响Fig.6 Effects of high temperature and its combined treatmentwith Na2SeO3 on total antioxidant capacity of broccoli sprouts
2.3 高温及其联合Na2SeO3处理对西兰花芽苗异硫氰酸酯代谢指标的影响
2.3.1 ITCs含量 发芽期间各处理下西兰花芽苗中ITCs含量均随发芽时间延长呈下降趋势,见图7。发芽4、6 d时,高温胁迫及其联合Na2SeO3处理的芽苗中ITCs含量较对照处理均显著提高(P<0.05),发芽4 d时,其ITCs含量分别是空白对照的2.36倍和3.35倍。同时,相较于单独高温胁迫处理,高温联合Na2SeO3处理的芽苗中ITCs含量也显著提高(P<0.05)。发芽6 d时,高温及其联合Na2SeO3处理的芽苗中ITCs含量分别是对照的2.1倍和2.46倍。研究结果表明,发芽4、6 d时,西兰花芽苗经高温及高温联合Na2SeO3处理均较空白对照可积累ITCs含量,且高温联合Na2SeO3处理4 d时,芽苗中ITCs含量最高达267 mg/100 g FW。
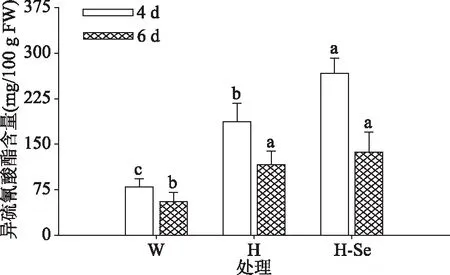
图7 高温及其联合Na2SeO3处理对西兰花芽苗异硫氰酸酯含量的影响Fig.7 Effects of high temperature and its combined treatmentwith Na2SeO3 on the content of isothiocyanate in broccoli sprouts
2.3.2 硫苷含量 如图8,各处理下西兰花芽苗中硫苷含量均随发芽时间显著降低(P<0.05)。发芽4 d时,对照和高温胁迫处理的芽苗中硫苷含量无显著变化(P>0.05),而高温联合Na2SeO3处理显著提高硫苷含量(P<0.05)。西兰花芽苗经高温及其联合Na2SeO3处理6 d,各处理硫苷含量无显著差异(P>0.05)。结果显示西兰花芽苗经高温联合Na2SeO3处理发芽4 d时效果最好,其芽苗中ITCs含量和硫苷含量均最高。

图8 高温及其联合Na2SeO3处理对西兰花芽苗硫苷含量的影响Fig.8 Effects of high temperature and its combined treatmentwith Na2SeO3 on glucosinolate content in broccoli sprouts
2.3.3 黑介子酶活性 由图9可看出,西兰花经高温联合Na2SeO3处理期间,其芽苗中MYR活性与对照无显著差异(P>0.05)。而高温胁迫处理下西兰花芽苗中MYR活性均显著低于对照处理(P<0.05);发芽4 d时,高温联合Na2SeO3处理的西兰花芽苗中MYR活性较高温胁迫处理无显著变化(P>0.05),而发芽6 d时,其MYR活性显著提高(P<0.05),说明高温联合Na2SeO3处理后对西兰花芽苗中MYR活性无显著影响。

图9 高温及其联合Na2SeO3处理对西兰花芽苗黑介子酶活性的影响Fig.9 Effects of high temperature and its combined treatmentwith Na2SeO3 on the activity of myrosinase in broccoli sprouts
3 讨论
植物在高温、低氧、盐害等逆境条件下,其生物膜受到损伤,同时细胞内活性氧代谢平衡失调,进而影响其蛋白质等生物大分子正常代谢,植物的生长和发育受到抑制。一般陆生高等植物热害界限温度为35 ℃,本研究中,西兰花发芽期间经高温(60 ℃)及其联合Na2SeO3处理后,其生命活力受到胁迫抑制,相较正常生长芽苗,胁迫处理下的植株形态较小,芽长显著降低。非生物胁迫下,植物体内积累ROS是一种普遍现象,其对植物细胞膜系统造成氧化伤害[20]。MDA 是细胞膜脂过氧化产物,其含量反映逆境下植物细胞的完整性[21]。西兰花芽苗经高温联合Na2SeO3处理,芽苗中 MDA和H2O2含量较对照均显著增加(P<0.05),表明高温联合Na2SeO3处理致使西兰花芽苗中活性氧累积,细胞膜脂过氧化严重。Peas等[22]发现适宜浓度硒处理可提高西兰花芽苗抗氧化能力,而Yang等[23]研究显示60 ℃热激时甘蓝的抗氧化能力达到最高,本研究同样发现西兰花芽苗经高温及其联合Na2SeO3处理后,发芽4 d时其总抗氧化能力较对照均显著提高,且其总抗能力高于发芽6 d。
非生物胁迫,如干旱、盐害和低温等均能够引起植物损伤,并诱导次生代谢产物的增加[24]。ITCs作为植物中具有抗癌活性的次生代谢产物而一直受到食品科研工作者的关注。本研究结果显示,西兰花发芽期间经高温及其联合Na2SeO3处理,发芽4 d和6 d时其ITCs含量均较对照处理显著提高,同时,高温联合Na2SeO3处理显著高于单独高温胁迫处理。Kim等[13]研究也表明利用Na2SeO3对盐胁迫下的甘蓝进行处理可有效提高其ITCs含量,这表明Na2SeO3联合非生物胁迫(高温胁迫和盐胁迫)具有单独环境胁迫的叠加作用,西兰花芽苗经高温联合Na2SeO3处理是富集ITCs的有效方式。ITCs的前体物质硫苷在不同处理下其含量同样发生显著变化,高温联合Na2SeO3处理4 d时,芽苗中硫苷含量较空白对照与高温胁迫处理均显著提高,而对照与高温胁迫处理则无显著差异,说明Na2SeO3处理显著提高芽苗中硫苷含量,然而随发芽时间延长,发芽6 d时西兰花芽苗中硫苷含量显著降低,且各处理间无显著差异,这与vila等[25]的研究结果相一致。MYR作为水解硫苷产生ITCs的关键酶,本研究中高温联合Na2SeO3处理发芽期间,西兰花芽苗中MYR活性均与对照处理无显著差异,且单独高温胁迫处理时其MYR活性显著低于对照处理。同样的,NaCl和CaCl2处理对西兰花芽苗中黑介子酶活性亦无显著影响[26]。我们推测高温及其联合Na2SeO3处理提高ITCs含量可能并非通过直接提高MYR活性达成,其可能与MYR以及ITCs合成途径中其他关键酶的转录水平有关,这有待后续深入研究高温及其Na2SeO3处理下西兰花芽苗中相关基因转录表达变化。
4 结论
西兰花芽苗发芽期间经高温胁迫及其联合Na2SeO3处理,其生长发育受到抑制,芽长显著降低,丙二醛含量显著增加,芽苗总抗氧化能力也显著增加。高温联合Na2SeO3处理下芽苗中ITCs含量及硫苷含量显著提高,高温处理下MYR活性显著降低。西兰花芽苗经高温联合Na2SeO3处理4 d,其ITCs和硫苷含量最高,表明高温联合Na2SeO3处理是富集西兰花芽苗中ITCs和硫苷的有效方式,且发芽时间以4 d为宜。
猜你喜欢
粘接(2022年12期)2023-01-05
畜牧兽医杂志(2018年2期)2018-06-15
现代园艺(2017年21期)2018-01-03
青年文学家(2017年28期)2017-11-28
江苏农业科学(2016年8期)2017-02-15
分析科学学报(2016年6期)2016-10-16
中国塑料(2016年4期)2016-06-27
食品与生活(2014年2期)2014-03-04
北京航空航天大学学报(2012年5期)2012-12-19
植物营养与肥料学报(2010年6期)2010-10-26
