
新昌宫廷黄鸡遗传多样性及遗传结构分析
2019-11-27 08:47:52王珍珍黄玲玲赵婉秋李志梁刘银兰詹海琴石孟达卢立志
中国畜牧杂志 2019年11期
王珍珍,黄玲玲,赵婉秋,李志梁,刘银兰,詹海琴,石孟达,田 勇,曾 涛,卢立志*
(1.浙江师范大学化学与生命科学学院,浙江金华 321001;2.浙江省农业科学院畜牧兽医研究所,浙江杭州 310023;3.福建农林大学动物科学学院,福建福州 350002;4.新昌县宫廷黄鸡繁育有限公司,浙江绍兴 312530)
微卫星是一种简单重复的2~4 个核苷酸序列基序,两侧有独特的序列,由于它们是共显性的,可以检测到高水平的等位基因多样性,并且易于用PCR 进行分析,因此作为遗传标记是有价值的[1]。微卫星标记,又称STR(Short Tandem Repeat)或SSR(Simple Sequence Repeats),通常用于研究自然种群的遗传结构,关于遗传变异在种群间的分配不仅在进化生物学和生态学中具有重要意义,而且在保护生物学中也具有重要意义[2]。由于其重复的特征,在DNA 复制过程中易发生DNA聚合酶“打滑”现象,导致重复序列中核心序列重复次数的改变而出现突变,因此STRs 也是基因组中变异最快的位点之一[3],所以STRs 通常与遗传疾病和基因调控功能有关,也是分析进化、遗传多样性和法医鉴定的遗传标记[4]。
我国是家禽饲养及消费大国,鸡更是生活中最常见的家禽之一,研究鸡的遗传多样性及遗传结构有助于对其进行合理的开发与利用。宫廷黄鸡是我国颇具特色的地方品种,具有独特的外貌特征和很高的观赏价值,蛋品质优良,肉质浓郁[5]。本研究利用微卫星对宫廷黄鸡遗传多样性进行检测,并分析其与浙江省不同地方鸡品种间的遗传结构,为今后新昌宫廷黄鸡种质资源的保护和合理的开发利用提供科学的理论支撑。
1 材料与方法
1.1 样品采集及DNA 提取 60 只宫廷黄鸡来自新昌县宫廷黄鸡繁育有限公司,其他5 个鸡品种为丝羽乌骨鸡(浙江合兴禽业发展有限公司)、龙游麻鸡(龙游县田野山庄)、萧山鸡(杭州萧山东海养殖有限公司)、仙居鸡(仙居鸡开发总公司)、白耳黄鸡(江山蓝丰种禽有限公司),各60 只。所有个体用含有抗凝剂Na2EDTA 或K2EDTA 的抗凝管进行翅下静脉釆血,低温保存,使用TaKaRa Blood Genome DNA Extraction Kit 提取基因组DNA。
1.2 引物信息及PCR 扩增 首先,根据GenBank 中所提供的鸡的SSR 序列利用Primer Premier 5 软件进行引物设计或者对相关文献中引物进行筛选,最终筛选出核心重复序列较多,且多态性丰富的位点。共有26 对引物扩增成功:ADL136,DL166,ADL185,ADL195,ADL210,ADL212,ADL225,ADL123,ADL176,LEI 0166,LEI 0066,MCW 0295,MCW 0014,MCW 67,MCW 0081,MCW 183,MCW 330,MCW 0085,MCW 264,MCW 134,MCW 104,MCW150,MCW32,MCW4,MCW120,MCW174,其 中ADL166、ADL185、MCW0014 和MCW0085 引物序列参考沈立权[6],MCW264 和MCW4 引物序列参考李红霞[7],ADL176 引物序列参考陈红菊等[8],引物由上海英捷生物有限公司合成。
将提取到的基因组DNA 以及合成的引物进行PCR反应,得到的PCR 产物于4℃保存备用,含荧光标记的PCR 产物用 ABI-3730XL DNA Analyzer 全自动测序仪检测DNA。PCR 反应体系、扩增反应程序以及检测程序参照陶争荣等[9]和刘雅丽等[10]。
1.3 统计分析 利用POPGENE 1.31 软件进行遗传多样性参数统计,利用GENEPOP 4.2[11]软件卡方检验HWE无偏估计概率值(P);利用Dispan 软件统计各群体间Nei 遗传距离DA[12],并进行邻接法(Neighbor-Joining,NJ)聚类分析,基于群体间遗传距离参数用MEGA 4.0[13]软件,构建品种间的系统发生树;运用Gentix4.02[14]软件中相关因子分析基于鸡群个体遗传差异进行成分因子分析。
2 结果与分析
2.1 遗传多样性分析 经统计(表1),去除ADL136、ADL195、ADL210 这3 个无效等位基因座(文中未呈现具体检验过程)。宫廷黄鸡26 个微卫星座平均等位基因数(Ne)为4.03 个,在座位MCW32 上检测到的基因数最多,在座位LEI0166 上检测到的基因数最少。宫廷黄鸡26 个微卫星座多态信息含量(PIC)均值为0.67,大部分位点PIC 大于0.5,部分位点(ADL123、LEI0166、MCW0085)介于0.25~0.5,为中等多态性微卫星座。观察杂合度(Ho)与期望杂合度(He)均值分别为0.60、0.72,平均Shannon 信息指数(I)为1.50。利用GENEPOP 进行 Hardy-Weinberg 平衡检验发现,经过邦佛伦尼[15]校正(Bonferroni correction)(P<0.002),26 个位点中有11 个位点处于不平衡状态。
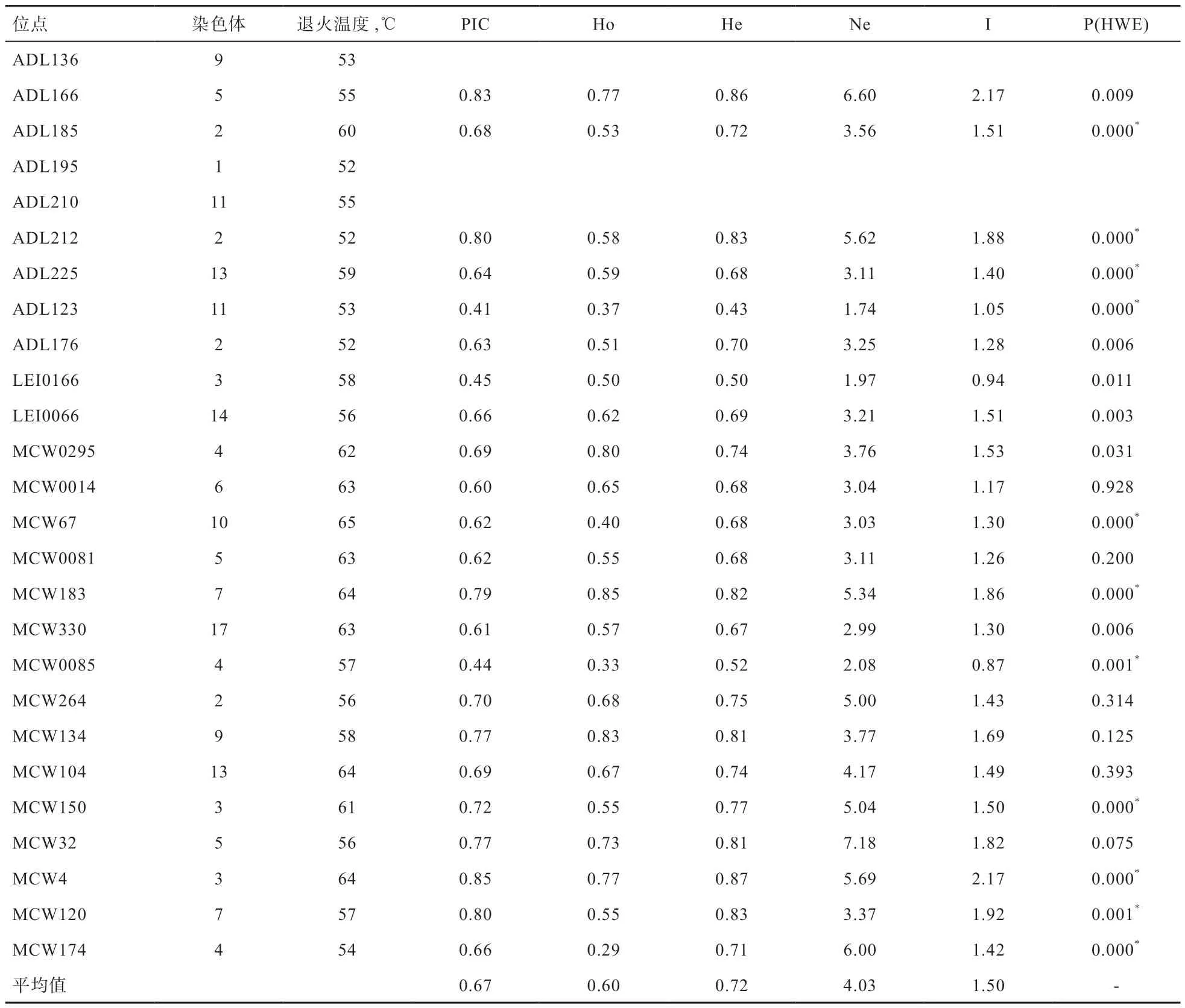
表1 26 对STR 引物信息以及遗传多样性参数
2.2 遗传距离估计 宫廷黄鸡与其他鸡品种间的遗传距离DA见表2,6 个鸡群体中仙居鸡和龙游麻鸡的DA遗传距离最小,仅为0.103 5,丝羽乌骨鸡和龙游麻鸡的DA遗传距离也比较小,为0.134 5,而宫廷黄鸡与其他群体之间的DA遗传距离均在0.190 0 以上,其中与丝羽乌骨鸡DA遗传距离最小,为0.190 9,与白耳黄鸡的DA遗传距离最大,为0.342 7,事实上,白耳黄鸡与其他群体之间的遗传距离均在0.300 以上,比其他群体间任意2 个群体间的DA遗传距离都高。
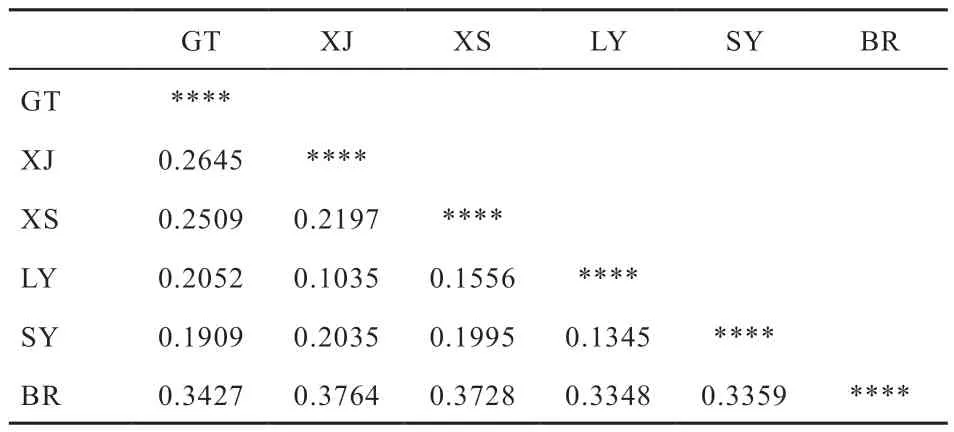
表2 宫廷黄鸡与其他品种鸡群DA 遗传距离
2.3 聚类分析 基于遗传距离DA采用NJ 构建系统发生树,结果见图1。宫廷黄鸡与其他4 个品种鸡群分为一簇,仙居鸡先与龙游麻鸡聚合再与萧山鸡、丝羽乌骨鸡聚合,最终与宫廷黄鸡聚合,而白耳黄鸡则单独作为一簇。
2.4 因子相关分析(FCA) 基于个体遗传差异的FCA 结果显示,反映个体间遗传差异最主要的3 个因子(Factor I=37.43%,Factor II=22.37%,Factor III=18.27%)呈现了总变异的78.08%,因而能较好反映群体簇聚情况。从FCA 结果可知,宫廷黄鸡依次与丝羽乌骨鸡、龙游麻鸡、萧山鸡、仙居鸡簇在一起,白耳黄鸡单独为一簇。
3 讨 论

图1 基于DA 遗传距离构建的系统发生树(NJ)
3.1 遗传多样性 遗传多样性是世界自然保护联盟(IUCN)公认的值得保护的3 种生物多样性形式之一。保护种群内遗传多样性需要基于2 个论点:进化发生的遗传多样性的必要性,以及杂合度与种群适应性之间的预期关系[16]。本研究采用26 个微卫星基因座,对宫廷黄鸡60 个个体进行了遗传多样性分析,发现宫廷黄鸡的26 个位点的平均PIC 为0.67,其中只有ADL123、LEI0166、MCW0085 3 个位点为中度多态(0.25<PIC<0.5),呈现出丰富的多态性;本研究所选用的26 个微卫星位点的平均杂合度高于0.60,为高度多态位点,表明宫廷黄鸡具有丰富的遗传多样性,而其Ho(0.60)低于He(0.72),说明其纯合子比较多,外界的变化对于其影响较小;Shannon 信息指数越大,群体遗传结构变异性越高,宫廷黄鸡26 个微卫星座的平均Shannon 信息指数I 为1.50,说明该群体遗传结构变异性比较高。哈代-温伯格平衡结果显示,在26 个位点中有11 个位点处于不平衡状态,说明这11 个位点的基因频率在后代中会发生改变。
3.2 遗传距离与系统发生树 遗传距离是衡量2 个不同种群或亲缘关系较近的物种之间遗传差异的一种方法,通常是利用2 个种群不同位点的等位基因频率数据来计算[17]。遗传距离测量是种群或物种间亲缘关系的指标,对重建种群间的历史和系统发育关系具有重要意义[18]。遗传距离的大小可以反映出群体间的亲缘关系[19-21]。本研究结果表明,与另外5 个品种相比较,宫廷黄鸡与丝羽乌骨鸡遗传距离最小,即亲缘关系最近,与白耳黄鸡亲缘关系最远;仙居鸡与萧山鸡的遗传距离大于仙居鸡与龙游麻鸡的遗传距离,这一结果与曾涛等[22]研究结果一致。总体来看,仙居鸡与龙游麻鸡的遗传距离最小,其次是仙居鸡与丝羽乌骨鸡,白耳黄鸡与其他群体之间的距离均在0.33 以上,属亲缘关系最远。以DA遗传距离为基础绘制了NJ 遗传进化树,宫廷黄鸡与其他4 品种鸡群分为一簇,仙居鸡先与龙游麻鸡聚合再与萧山鸡、丝羽乌骨鸡聚合,最终与宫廷黄鸡聚合,而白耳黄鸡则单独作为一簇,聚类结果与DA遗传距离反映结果一致。
3.3 FCA FCA 是对数据集的归纳探索,通过将主要性状的数量减少到更少的因子中,这些因子是原始变量的组合,是方差最大的因子,消除了原始数据集中的冗余,可以发现性状之间的真实关联[23]。本研究结果显示,6个不同鸡群间的遗传结构,与基于遗传距离NJ 系统树分析结果一致,白耳黄鸡单独为一簇,宫廷黄鸡与丝羽乌骨鸡最近,而龙游麻鸡与仙居鸡及萧山鸡遗传距离较近。
4 结 论
本研究利用 26 个微卫星位点分析宫廷黄鸡群体遗传多样性及遗传结构的结果显示,新昌宫廷黄鸡具有丰富的遗传多样性,具有较高的保种价值;遗传距离分析和聚类分析显示,宫廷黄鸡与丝羽乌骨鸡遗传距离最近,与白耳黄鸡遗传距离最远。
猜你喜欢
艺术家(2024年1期)2024-03-02 08:01:22
小猕猴学习画刊·下半月(2023年4期)2023-06-20 19:29:14
今日农业(2022年13期)2022-09-15 01:18:40
科普童话·学霸日记(2021年3期)2021-09-05 16:42:08
中国畜禽种业(2020年7期)2020-12-16 14:47:06
国学(2020年1期)2020-06-29 15:14:36
作文小学中年级(2019年10期)2019-11-04 00:39:52
幼儿100(2017年13期)2017-06-15 20:29:45
中国畜牧兽医文摘(2016年7期)2016-01-30 23:38:24
中国记者(2014年1期)2014-03-01 01:37:37
