
利用KASP标记检测青海和西藏小麦品种中光周期基因分布
2019-11-22 07:10权有娟袁飞敏刘德梅陈志国
麦类作物学报 2019年10期
权有娟,袁飞敏,刘德梅,李 想,陈志国
(1.中国科学院西北高原生物研究所/中国科学院高原生物适应与进化重点实验室/青海省作物分子育种重点实验室,青海西宁 810008; 2.中国科学院大学,北京 100049; 3.西北农林科技大学,陕西杨凌 712100)
小麦是世界上第三大粮食作物,也是我国的主要粮食作物。普通小麦对不同环境的适应性受开花时间影响较大[1],主要由3种基因决定,分别是春化基因(Vrn)、光周期基因(Ppd)和早熟性基因(Eps)[2],前两者易受环境的影响,后者不受环境影响。其中,光周期反应主要由3个同源主效基因控制,即Ppd-D1(Ppd1)、Ppd-B1(Ppd2)和Ppd-A1(Ppd3),分别位于小麦2D、2B和2A染色体上[3-4]。只有在长日照条件下才能开花的小麦为光周期敏感类型,在长日照或短日照条件下均可开花的小麦为光周期不敏感类型[5]。通常认为等位变异Ppd-D1a对光周期反应最不敏感,其次是Ppd-B1a、Ppd-A1a[8],但Tanio等[9]证实Ppd-B1a和Ppd-D1a的光周期效应相同。目前鉴定的小麦光周期基因均有光周期不敏感型等位变异Ppd-D1a、Ppd-B1a、Ppd-A1a和光周期敏感型等位变异Ppd-D1b、Ppd-B1b、Ppd-A1b两种等位变异类型[6-7]。光周期基因不仅影响着小麦的生育期和产量,而且对小麦的适应性、种质资源创制、新品种引种和扩大种植范围等极其重要[10]。
功能标记在作物育种过程中发挥着重要作用,在小麦中已获得100多个基因的功能标记,为育种家提供了一个对有利性状选择的分子工具箱[11]。Beales等[12]利用基因序列特异性开发了小麦光周期基因的功能标记,检测出Ppd-D1b和Ppd-D1a两种等位变异,Nishida等[13]利用同样的原理检测出了Ppd-B1和Ppd-A1的等位变异。KASP(kompetitive allele specific PCR)标记是近几年在作物分子标记辅助与选择育种中常用的方法,该标记技术具有高通量辅助筛选功能,使快速提高小麦育种效率和改良品种成为可能。Rasheed等[14]开发和鉴定了一些与普通小麦产量、品质、适应性、抗性有关的KASP标记,证明KSAP标记的检测速度是基于凝胶PCR分子标记的45倍。Semagn[15]等研究发现,KASP标记和传统的SNP标记检测技术相比,具有更高效、费用低、更精确的特点。
青藏高原是我国小麦高产区,但品种多在本地区表现适宜性好、高产,离开青藏高原往往表现不佳,难以适应。对其光周期反应加以研究,可以有助于了解高原品种的生态适应性。国内外对光周期基因Ppd-D1的组成和分布研究较多,对Ppd-B1和Ppd-A1基因的研究较少。青藏高原地形复杂多样,小麦类型丰富,对青海和西藏地区小麦品种光周期基因的组成和分布开展研究,可为今后在这一地区培育广适小麦品种以及新品种引种、推广提供理论理论支持和依据。
1 材料与方法
1.1 材 料
供试小麦品种(系)共计249份,包括70份青海省历年审定品种、75份外引资源品种(系)、27份自育高代品系、42份青海农家品种及35份西藏农家品种,详细信息见表1。
1.2 方 法
1.2.1 基因组DNA提取
供试品种(系)各挑选种子30粒,室温条件下发芽,采用CTAB法[16]提取植物基因组总DNA,利用1%琼脂糖凝胶电泳和紫外分光光度计检测DNA纯度和浓度。
1.2.2 KASP标记检测
实验所需的KSAP标记引物由上海生工股份有限公司合成(表2),引物序列参考Rasheed等[14]。FAM 5′端的标签序列为:5′-GAAGGTG ACCAAGTTCATGCT-3′,VIC 5′端的标签序列为:5′-GAAGGTCGGAGTCAACGGATT-3′。共需3组混合引物,每组混合引物由2条各12 μL(100 μmol·L-1)的正向序列、1条的30 μL(100 μmol·L-1)反向共同序列及46 μL ddH2O组成。

表1 249份供试材料的名称Table 1 Sources andnames of the 249 test materials
(续表1 Continued table 1)

序号 Number品种(系)名称 Variety(line) name序号 Number品种(系)名称 Variety(line) name序号 Number品种(系)名称 Variety(line) name45青农469 Qingnong 469128济麦262 Jimai 262211兰麦-2 Lanmai-246宁春26号 Ningchun 26129德抗961 Dekang 961212火麦 Huomai47互麦12号 Humai 12130晋2148 Jin 2148213和尚麦 Heshangmai48香农3号 Xiangnong 3131太空8号 Taikong 8214青海北 Qinghaibei49高原392 Gaoyuan 392132京冬8号 Jingdong 8215怒江小麦 Nujiangxiaomai50青春587 Qingchun 587133京冬16 Jingdong 16216曲水小麦 Qushuixiaomai51高原466 Gaoyuan 46613403中35 03 zhong 35217红颖无芒 Hongyingwumang52互麦14号 Humai 1413507中38 07 zhong 38218卓果索 Zhuoguosuo53乐麦6号 Lemai 6136陕农69 Shaannong 69219短密穗长芒 Duanmisuichangmang54乐麦5号 Lemai 5137云麦53 Yunmai 53220半芒春麦 Banmangchunmai55通麦1号 Tongmai 1138辽春10号 Liaochun 10221贡嘎小麦 Gonggaxiaoami56青春6号 Qingchun 6139冀麦586 Jimai 586222早熟小麦 Zaoshuxiaomai57青春10号 Qingchun 10140绵阳096-61 Mianyang 096-61223宋噶小麦 Songgaxiaomai58墨引2号 Moyin 21418616M7224扎仁卓玛 Zharenzhuoma59青加2号 Qingjia 2142新旱212 Xinhan 212225扎西布索 zhaxibusuo60高原671 Gaoyuan 671143新旱688 Xinhan 688226白小麦 Baixiaomai61互麦15号 Humai 1514411MJ557227扎其毛颖 Zhaqimaoying62青加1号 Qingjia 1145关东107 Guandong 107228柳乡呸卓 Liuxiangpeizhuo63青春592 Qingchun 5921469097229古曲扎通 Guquzhatong64紫糯1号 Zinuo 114717478230恰噶小麦 Qiagaxiaomai65曹选3号 Caoxuan 314814609-24231丁青基卓 Dingqingjizhuo66新哲9号 Xinzhe 914914609-1232山南大碎粒 Shannandasuili67烟幅188 Yanfu 18815014609-2233仁达代光麦 Rendadaiguangmai68兰考906 Lankao 90615114609-3234纹龙小麦 Wenlongxiaomai69马品 Mapin1529099235扎西才卓玛 Zhaxicaizhuoma70青春593 Qingchun 59315308654236昌都小麦 Changduxiaomai71甘麦8号 Ganmai 81549096237卓鲁娜 Zhuoluna72定西24 Dingxi 241552109238早红芒麦 Zaohongmangmai73甘麦35 Ganmai 351569101239左贡圆锥麦 Zuogongyuanzhuimai74陇春17 longchun 1715791771240白龙春麦 Bailongchunmai75陇春13 Longchun 13158121-4241变芒麦 Bianmangmai76甘春20 Ganchun 20159074242彼巴密穗 Bibamisui77陇春2609 Longchun 2609160056243隆子圆锥 Longziyuanzhui78张811 Zhang 811161052244尼木密穗 Nimumisui79武春4号 Wuchun 416290-1073245白当卓尔 Baidangzhuoer80兰天3号 Lantian 316314313246卧龙波兰 Wolongbolan81张春6号 Zhangchun 61649106247斗玉波兰 Douyubolan82武春2号 Wuchun 21659161248洋松 Yangsong83甘春358786 Ganchun 3587861668337249加察碎穗麦 Jiachasuisuimai
1~70号为青海育成品种:71~145号为青海引进品种(系);146~172号为青海高代品系;173~214号为青海农家品种;215~249号为西藏农家品种。
1-70 are bred varieties of Qinghai; 71-145 are introduced varieties(line) of Qinghai; 146-172 are high-generation lines of Qinghai; 173-214 are Qinghai landraces of Qinghai; 215-249 are landraces of Tibet.
检测光周期基因等位基因型的PCR反应体系为5 μL,总体积中含2.0 μL DNA(10 ng·μL-1),2.5 μL KSAP Master Mix(LGC Genomics,Hoddeston,UK)、0.07 μL KSAP及 0.43 μL ddH2O。设置用ddH2O 代替模板DNA的空白对照,以防对试验结果造成误判。PCR在S1000TMThermal Cycler PCR仪上进行。PCR反应程序为:95 ℃,15 min;95 ℃变性 20 s,65 ℃退火和延伸25 s,57 ℃退火和延伸60 s,30个循环。反应结束后进行荧光扫描和基因分型分析。

表2 光周期基因的KASP标记引物Table 2 KASP primers used in this study for photoperiod genes
该表中横杠不代表引物序列缺失,而表示TaPpdBJ003中没有FAM引物序列。
The dotted line in this table indicates there is no FAM primer sequence in TaPpdBJ003.
2 结果与分析
2.1 光周期基因位点等位变异的KASP检测 结果
2.1.1Ppd-D1位点鉴定结果
利用TaPpdDD001、TaPpdDI001和 TaPpdDD002引物组合对供试材料进行Ppd-D1基因位点的等位变异检测。结果(表3及其注释)显示,青海的42份农家品种中,红农1号、大庄火麦、群科大白麦等41份材料中检测出光周期不敏感型等位变异Ppd-D1a,仅有兰麦-2含有光周期敏感型等位变异Ppd-D1b;75份外引资源中,甘春20、巴丰5号、宁春4号等64份材料检测到含有光周期不敏感型等位变异Ppd-D1a,晋2148、关东107、内麦18等11份材料检测出了光周期敏感型等位变异Ppd-D1b;70份育成品种中,柴春044、柴春901、高原448等60份材料检测出光周期不敏感型等位变异Ppd-D1a,烟幅188、高原602、乐麦6号等10份材料检测出光周期敏感等位变异Ppd-D1b;27份高代品系中,24份材料检测出光周期不敏感型等位变异Ppd-D1a,056、08654、97-2051检测出光周期敏感型等位变异Ppd-D1b。在35份西藏农家品种中,尼木密穗、昌都小麦、曲水小麦等32份材料检测出光周期不敏感型等位变异Ppd-D1a,彼巴密穗、怒江小麦、纹龙小麦检测出光周期敏感型等位变异Ppd-D1b。
2.1.2Ppd-B1位点的鉴定结果
用TaPpdBJ001、TaPpdBJ003引物组合对供试材料进行Ppd-B1基因位点的等位变异检测,结果(表3及其注释)表明,青海42份农家品种、75份引进品种、27份高代品系都只携带光周期不敏感型等位变异Ppd-B1a;70份育成品种中,只有高原314携带光周期敏感型等位变异Ppd-B1b,其余材料均为光周期不敏感型Ppd-B1a。西藏的35份农家品种中,卓果索、宋噶小麦、卓鲁娜等24份材料携带光周期不敏感型等位变异Ppd-B1a,仁达代光麦、山南大碎粒、恰噶小麦等11份材料携带光周期敏感型等位变异Ppd-B1b。
2.1.3Ppd-A1位点的鉴定结果
引物组合Cdex5-6ID、GS100-1027ID和GS105-1117ID在Ppd-A1基因位点的等位变异检测结果(表3及其注释)表明,青海的42份农家品种中,仅有大红头麦携带光周期敏感型等位变异Ppd-A1b,其余品种均携带光周期不敏感型等位变异Ppd-A1a;75份引进品种中,济麦17号、兰考906、良星99等67份材料携带光周期不敏感型等位变异Ppd-A1a,张春6号、甘麦35、武科2246等8份材料携带光周期敏感型等位变异Ppd-A1b;70份育成品种中,青春570、青春587、墨引2号等63份携带光周期不敏感型等位变异Ppd-A1a;高原V028、辐射阿勃、乐麦5号等7份材料携带光周期敏感型等位变异Ppd-A1b;27份高代品系均携带光周期不敏感型等位变异Ppd-A1a。西藏的35份农家品种均携带光周期不敏感型等位变异Ppd-A1a。
2.2 光周期基因等位变异的分布
等位变异在249份材料中的平均分布频率不同,3个光周期不敏感型等位变异中,Ppd-B1a和Ppd-A1a出现的平均比例均在90%以上,Ppd-D1a所占比例稍低一点,为88.76%。3个光周期敏感型等位变异中,Ppd-D1b出现的平均比例最高(占11.24%),Ppd-A1b和Ppd-B1b的平均分布频率均在10%以下(表3)。
青海不同类型小麦品种中,以光周期不敏感型等位变异Ppd-B1a为主,出现的频率均在98%以上,其中,高代品系中还以光周期不敏感型等位变异Ppd-A1a为主(占100%),光周期敏感型等位变异出现的频率较少,最高为引进品种(系)中的Ppd-D1b(占14.67%)。青海和西藏小麦农家品种中光周期基因等位变异的分布也存在差异,青海农家品种以光周期不敏感型等位变异Ppd-B1a为主(占100%),西藏农家品种以光周期不敏感型等位变异Ppd-A1a为主(占100%);光周期敏感型等位变异Ppd-B1b在西藏农家品种中出现的频率最高(占31.42%),青海农家品种中均在10%以下。
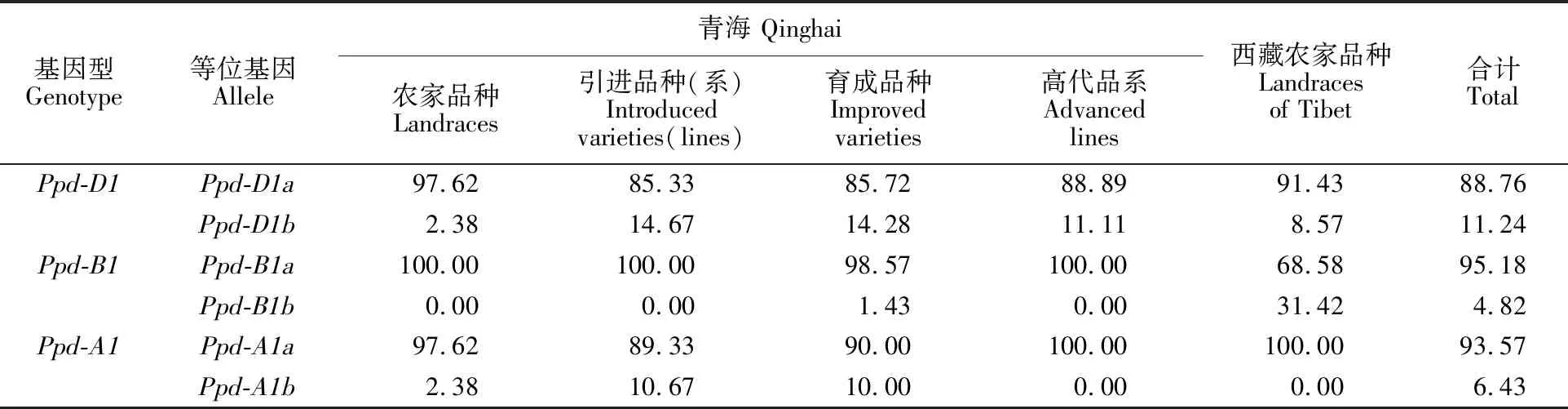
表3 光周期等位变异在青海和西藏小麦品种中的分布频率Table 3 Distribution of alleles at photoperiod gene loci among Qinghai and Tibet wheat cultivars %
1~5、7~8、10~11、13~23、25~26、29~47、49~50、52、54~66、68~72、76~84、86~115、117~118、120~125、127~129、131~136、138~139、141~144、146~152、154~159、161~171、173~210、212~214、216~233、235~241、243~249号品种携带等位变异Ppd-D1a;6、9、12、24、27~28、48、51、53、67、73~75、85、116、119、126、130、137、140、145、153、160、172、211、215、234、242号品种携带等位变异Ppd-D1b;1~8、10~216、218~219、221、223~227、229、231、236~241、243~249号品种携带等位变异Ppd-B1a;9、217、220、222、228、230、232~235、242号品种携带等位变异Ppd-B1b;1~13、15~17、19~32、35~38、 40~52、54~61、63~72、76~80、82~88、91~120、122~125、127~205、207~249号品种携带等位变异Ppd-A1a;14、18、33~34、39、53、62、73~75、81、89~90、121、126、206号品种携带等位变异Ppd-A1b。
1-5, 7-8, 10-11, 13-23, 25-26, 29-47, 49-50, 52, 54-66, 68-72, 76-84, 86-115, 117-118, 120-125, 127-129, 131-136, 138-139, 141-144, 146-152, 154-159, 161-171, 173-210, 212-214, 216-233, 235-241, and 243-249 varieties carry the allelic variationPpd-D1a;6, 9, 12, 24, 27-28, 48, 51, 53, 67, 73-75, 85, 116, 119, 126, 130, 137, 140, 145, 153, 160, 172, 211, 215, 234, and 242 varieties carry the allelic variationPpd-D1b;1-8, 10-216, 218-219, 221, 223-227, 229, 231, 236-241, and 243-249 varieties carry the allelic variationPpd-B1a; 9, 217, 220, 222, 228, 230, 232-235, and 242 varieties carry the allelic variationPpd-B1b; 1-13, 15-17, 19-32, 35-38, 40-52, 54-61, 63-72, 76-80, 82-88, 91-120, 122-125, 127-205, and 207-249 varieties carry the allelic variationPpd-A1a; 14, 18, 33-34, 39, 53, 62, 73-75, 81, 89-90, 121, 126, and 206 varieties carry the allelic variationPpd-A1b.
2.3 光周期基因等位变异组合的分布频率
青海和西藏小麦品种中共存在6种光周期基因等位变异组合类型,每种等位变异组合类型的平均分布频率不同。等位变异组合Ppd-D1a/Ppd-B1a/Ppd-A1a出现的频率最高,平均为 81.12%;其余几种类型的分布频率较低,都在10%以下(表4)。
青海小麦品种中存在5种等位变异组合类型,以Ppd-D1a/Ppd-B1a/Ppd-A1a类型为主(占 82.24%),其余类型的出现频率都在10%以下。青海和西藏农家品种中的等位变异组合类型也存在差异,青海农家品种中存在3种等位变异组合类型,以Ppd-D1a/Ppd-B1a/Ppd-A1a类型为主(占95.24%);西藏农家品种中存在4种等位变异组合类型,但也以Ppd-D1a/Ppd-B1a/Ppd-A1a类型为主(占65.71%)。
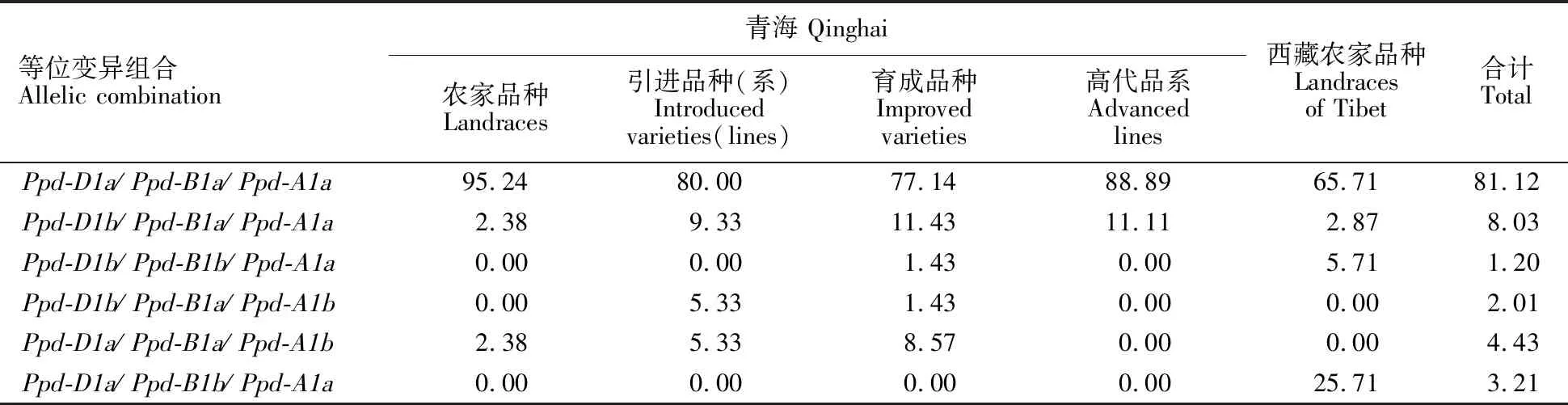
表4 光周期显性等位变异组合在青海不同麦区的分布比例Table 4 Distribution of dominant allele combinations at photoperiod gene loci in Qinghai and Tibet wheat zones %
1~5、7~8 、10~11、13、15~17、19~23、25~26、29~32、35~38、40~47、49~50、52、54~61、63~66、68~72、76~80、82~84、86~88、91~115、117~118、120、122~125、127~129、131~136、138~139、141~144、146~152、154~159、161~171、173~205、207~210、212~214、216、218~219、221、223~227、229、231、236~241、243~249号品种具有Ppd-D1a/Ppd-B1a/Ppd-A1a组合类型;6、12、24、27~28、48、51、67、85、116、119、130、137、140、145、153、160、172、211、215号品种具有Ppd-D1b/Ppd-B1a/Ppd-A1a组合类型;9、234、242号品种具有Ppd-D1b/Ppd-B1b/Ppd-A1a组合类型;53、73~75、126号品种具有Ppd-D1b/Ppd-B1a/Ppd-A1b组合类型;14、18、33~34、39、62、81、89~90、121、206号品种具有Ppd-D1a/Ppd-B1a/Ppd-A1b组合类型;217、220、222、228、230、232、233、235号品种具有Ppd-D1a/Ppd-B1b/Ppd-A1a组合类型。
1-5, 7-8, 10-11, 13, 15-17, 19-23, 25-26, 29-32, 35-38, 40-47, 49-50, 52, 54-61, 63-66, 68-72, 76-80, 82-84, 86-88, 91-115, 117-118, 120, 122-125, 127-129, 131-136, 138-139, 141-144, 146-152, 154-159, 161-171, 173-205, 207-210, 212-214, 216, 218-219, 221, 223-227, 229, 231, 236-241, and 243-249 varieties havePpd-D1a/Ppd-B1a/Ppd-A1acombination type; 6, 12, 24, 27-28, 48, 51, 67, 85, 116, 119, 130, 137, 140, 145, 153, 160, 172, 211, and 215 varieties havePpd-D1b/Ppd-B1a/Ppd-A1acombination type; 9, 234, 242 and varieties havePpd-D1b/Ppd-B1b/Ppd-A1acombination type; 53, 73-75, and 126 varieties havePpd-D1b/Ppd-B1a/Ppd-A1bcombination type; 14, 18, 33-34, 39, 62, 81, 89-90, 121, and 206 varieties havePpd-D1a/Ppd-B1a/Ppd-A1bcombination type; 217, 220, 222, 228, 230, 232, 233, and 235 varieties havePpd-D1a/Ppd-B1b/Ppd-A1acombination type.
3 讨 论
青海和西藏属于我国十大麦区之一的青藏春冬兼播麦区[17],虽属同一麦区,但两地的农家品种中光周期等位变异及组合类型分布存在一定差异。青海农家品种光周期不敏感型等位变异以Ppd-B1a占主导地位,Ppd-D1a和Ppd-A1a次之;西藏农家品种中光周期不敏感型等位变异以Ppd-A1a占主导地位,Ppd-D1a次之,Ppd-B1a最低。此研究结果与杨芳萍[18]、韩领锋等[19]的研究结果略有不同,可能与青海、西藏特殊的地理环境及气候有关,作物可以通过生理生化等变化协调自身的生长发育以适应不同的环境[20],因此,同一种质的基因和不同环境的互作导致植物产生不同的基因型和表现型。青海和西藏位于青藏高原腹地,纬度高、气温低、光照时间长,对光周期反应较敏感的小麦品种适应这种环境,以防止过早的形成花原基提前开花遭受霜冻[21]。两地农家品种共同拥有Ppd-D1a/Ppd-B1a/Ppd-A1a、Ppd-D1b/Ppd-B1a/Ppd-A1a组合类型,青海农家品种中还含有Ppd-D1a/Ppd-B1a/Ppd-A1b,西藏农家品种中另含Ppd-D1b/Ppd-B1b/Ppd-A1a和Ppd-D1a/Ppd-B1b/Ppd-A1a。两地农家品种中各自拥有的等位变异组合类型的品种在农艺形状上表现及与敏感性有何关系有待进一步研究。
光周期反应对品种的适应性有重要影响,本研究结果表明,随着种质资源的交流,越来越多的育成品种含有光周期不敏感基因,对光周期敏感性降低。只有在青海柴达木盆地超高产区(超长、超强光照条件)培育的品种(柴春901、高原465、高原466等),对生态条件具有特殊的要求,因此对光周期要求严格一些。
虽然多数品种以携带光周期不敏感型等位变异为主,无论是青海本土品种还是引进品种亦或是本土品种和引进品种杂交选育的育成品种及高代品系,光周期等位变异均以Ppd-B1a居多,推测这对青海育成品种的适应性有一定的限制。另外,生产实践表明,青海由于生态环境多样,地理条件和气候千差万别,为了保证一定产量,在育种中多选用高秆、大穗、高粒重材料,这些材料在我国其他麦区的气候条件很难得到满足,这也是高原小麦难以扩展的原因之一。本研究也推测出光周期反应可能不是影响青海和西藏小麦品种适应性强弱的主要原因,对于这一地区高产小麦品种难以大面积推广的原因还值得进一步深究。今后,青藏高原小麦育种中应该加强品种的引进,增加品种的多样性,以期提高小麦的适应性。
猜你喜欢
作物学报(2022年6期)2022-04-08
国际医学放射学杂志(2021年5期)2021-10-22
蔬菜(2020年4期)2020-12-14
党的生活·青海(2019年12期)2019-12-23
浙江农业学报(2019年7期)2019-08-14
金桥(2018年3期)2018-12-06
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
第一财经(2017年36期)2017-09-25
岷峨诗稿(2017年4期)2017-04-20
