
不同放牧强度对高寒草甸有毒植物多样性的影响
2019-11-19 05:59:34王志鹏张兆杰花立民
草原与草坪 2019年5期
王志鹏,张兆杰,花立民
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
青藏高原拥有我国乃至世界独特的高寒草甸生态系统,对中国和亚洲的生态安全具有重要影响[1]。近年来,在全球气候变化和人为干扰下,高寒草甸退化严重,毒杂草在草地植物群落中不断增加,禾本科、莎草科、豆科等可食牧草比例逐渐下降,对草地生产力和畜牧业的发展造成严重影响[2-5]。
放牧是天然草地最常见的利用方式[6],放牧干扰对草地植物群落以及多样性的影响已是草地生态学的研究热点之一。家畜通过选择性采食直接影响植物的种群动态,进而改变群落结构[7-8]。有毒植物生长速度快、繁殖力强等特点使其能够快速侵占大面积的天然草地,同时,有些有毒植物可以向环境中释放大量的化感物质,抑制牧草生长从而导致草地退化[9-10]。
近年来,国内外学者对放牧强度和植被群落特征的关系进行了较多的研究报道,但对放牧强度与有毒植物群落在时间尺度上的动态变化报道较少。基于此,以青藏高原东缘高寒草甸为研究对象,分析放牧强度与有毒植物物种多样性在放牧期内的关系,以明晰放牧对有毒植物的影响程度,为进一步研究有毒植物在草原生态系统生态和生产功能的作用和地位奠定基础。
1 材料和方法
1.1 研究区概况
研究区位于东祁连山的天祝金强河河谷,地理位置为N 37°11′48″,E 102°47′10″,海拔2 960 m。海拔高,气温低而寒冷,昼夜温差大,日照强烈,水热同期。年均气温-0.3~0℃,最冷月1月平均气温-18.3℃,最热月7月平均气温12.7℃,≥0℃年积温为1 300℃;年降水量416 mm,多集中在生长期;年蒸发量1 590 mm,约为年降水量的4倍。可食性牧草主要以矮嵩草(Kobresiahumilis)、西北针茅(Stipasareptana)、垂穗披碱草(Elymusnutans)、阴山扁蓿豆(Medicagoruthenica)、冷地早熟禾(Poaannua)为主。有毒植物主要有黄花棘豆(Oxytropisochrocephala)、高山唐松草(Thalictrumalpinum)、变异黄芪(Astragalusvariabilis)、秦艽(Gentianamacrophylla)等[11-12]。
1.2 样地设置
试验样地在天祝金强河甘肃农业大学试验站。放牧家畜为高山细毛羊。根据试验地多年来的地上植物生物量,放牧时间以及草地面积确定放牧强度,并以当地牧户近年来的放牧强度作为参考,在试验地设置轻度(LG)、中度(MG)、重度(HG)和过度(OG)4个放牧梯度(表1)。

表1 研究样地设置
1.3 野外调查与指标测定
于2015年6月中旬的牧草泛青期到10月中旬开展野外草地植物群落监测试验。采用随机固定W型路线的方法[12]在样地选取9个50 cm×50 cm的样方,测定各物种的盖度,过度放牧区由于面积较小,随机选取3个。在各样地随机抛面积为0.1 m2的样圆60次,测出现在的所有植物种的频度。每种植物测量株高10次。
1.4 数据分析
(1)物种丰富度指数(D),测定有毒植物物种数:
D=S
式中:S为样方内出现的有毒植物物种数。
(2)生物多样性指数,采用Shannon-Wienner指数(H):
H=-∑PilnPi
式中:Pi为种i的个体数在群落所有种中所占的重要值。
(3)均匀度指数,采用Pielou指数(E):
E=H/Hmax
式中:H为实际观察的物种丰富度指数,Hmax为最大物种丰富度性指数,Hmax= lnS,S为群落处理样地的总物种数。
采用SPASS 20.0软件进行不同放牧强度与有毒植物优势度、物种丰富度指数、生物多样性指数、均匀度指数的差异显著性分析和显著性检验。
2 结果与分析
2.1 试验区有毒植物的种类
根据《中国有毒植物》《甘肃草地资源》等文献资料,统计出试验区内所有有毒植物种类及名称。试验区出现有毒植物23种,分属于13科,以毛茛科、玄参科、菊科、豆科为主[13-14]。
2.2 有毒植物生物多样性在时空尺度上的变化
2.2.1 有毒植物丰富度变化 在各个特定的放牧强度下,有毒植物丰富度不同月季间出现重要变化。轻度放牧下,随时间的推移,8,9和10月之间出现显著差异,并且在8月达到最大值。中度放牧下,8、9月丰富度较高,显著高于其他月。在重度放牧下,有毒植物种数以7月为最高,依次为8、9月。在过度放牧区,8月有毒植物丰富度显著高于其他月,其余月份有毒植物丰富度随时间递减。在不同的月份,有毒植物丰富度在不同的放牧强度下呈现出明显变化。6月,在轻度放牧下达到最大,其次为过度放牧。7月,重牧区域内有毒植物丰富度都达到最大,其次为轻牧。8月,不同放牧强度下均出现显著差异,排序为轻度放牧>过度放牧>中度放牧>重度放牧。9月,轻度放牧下有毒植物丰富度出现最大值,而且中度放牧下的值也较大。10月,物种丰富度在放牧梯度下表现出的趋势与其他月不同,排序为重度放牧>中度放牧>轻度放牧>过度放牧。整体表现为有毒植物丰富度在7月或8月达到最高值,之后丰富度表现出下降的趋势。在轻度放牧和过度放牧强度下,有毒植物在6~8月丰富度高于中度放牧和重度放牧,在8月后低于中度放牧和重度放牧(表3)。

表2 天祝县抓喜秀龙高寒草甸试验区中有毒植物种类
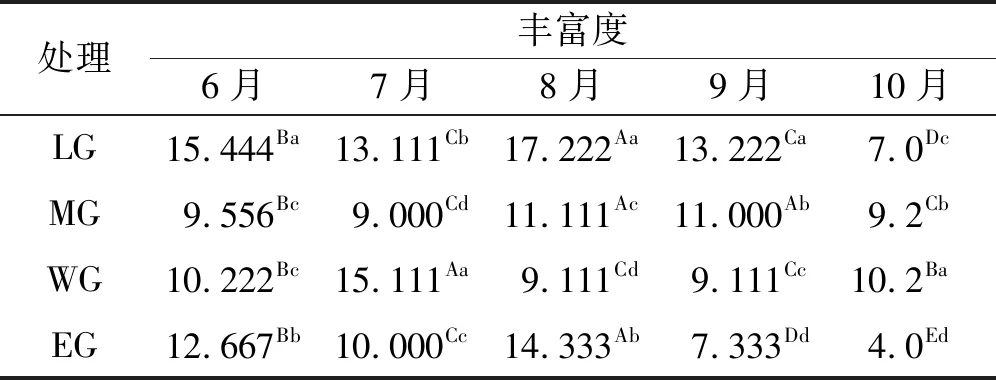
表3 有毒植物的物种丰富度
注:同行不同大写字母表示不同月份间差异显著(P<0.05);同列不同小写字母表示不同放牧强度下差异显著(P<0.05)。下同
2.2.2 有毒植物Shannon-Wiener指数变化 祁连山东段高寒草甸有毒植物Shannon-Wiener指数随着放牧强度的不同而有所变化(表4)。从时间尺度上分析,6月的有毒植物群落Shannon-Wiener指数较大,在轻度放牧达到最大值。在物种生育期末的9、10月,Shannon-Wiener指数显著低于前面的月份,并且除了重度放牧区,2个月之间均出现显著差异。从空间尺度分析,轻度放牧下,毒草的多样性指数较高,在6,8和9月达到最大值。不同月表现出不同的变化趋势,6月与8月的Shannon-Wiener指数从重度放牧到过重度放牧表现出升高的趋势。10月不同处理的的数值均显著小于其他月,值得注意的是,在重度放牧区多样性指数显著高于中度放牧区。过度放牧下,毒草的多样性指数在各月均低于轻度放牧,6月是过度放牧区毒草多样性指数最高的时间。10月过度放牧区毒草多样性指数要低于其他放牧处理。除重度放牧处理外,其他处理下,毒草的多样性指数都表现出7月降低、8月升高,之后的2个月降低的趋势。整体分析,试验区内有毒植物Shannon-Wiener 指数随植物生长季表现出7月分降低,8月分升高之后逐步降低的趋势,既使放牧强度不同,在10月都达到了Shannon-Wiener 指数最低值。
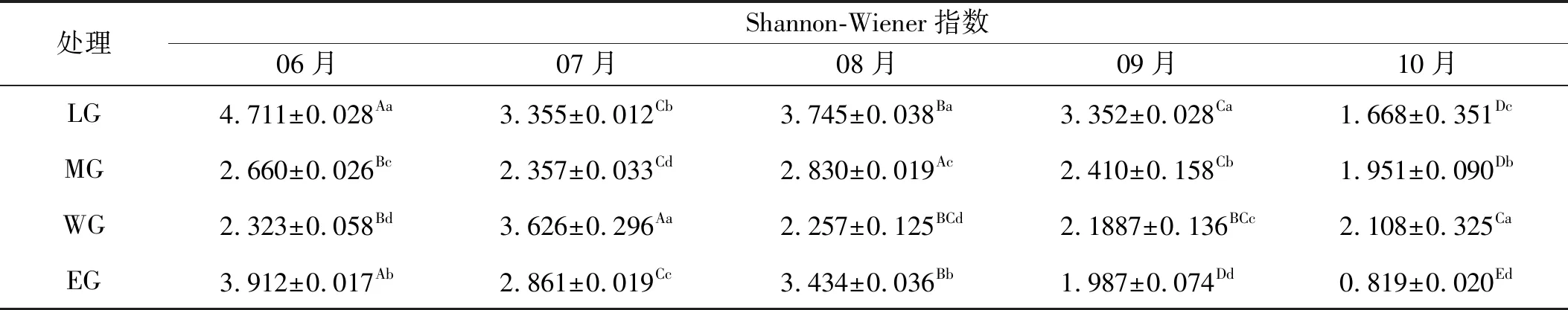
表4 试验区有毒植物的Shannon-Wiener指数
2.2.3 有毒植物Pielou指数变化 祁连山东段高寒草甸有毒植物Pielou指数随着生境的时空改变而有所变化(表5)。从时间尺度分析,轻度放牧下,Pielou指数月季间均出现显著差异,10月显著小于其他月。中度放牧下,表现出先降低后升高的趋势,6月Shannon-Wiener值较高,8,9和10月无显著差异。重度放牧下,返青期的Pielou指数较小,7月达到最大,10月Shannon-Wiener指数值也较高。过度放牧下,7、8月值较大,随时间的推移,表现出显著降低的趋势。从空间尺度分析,特定月份不同处理均表现出显著差异。6月中度放牧区的值最大,与其他各区差异显著。7月,不同处理间均为显著差异,重牧区最大,轻牧区值最小。8月,轻度放牧区达到最大值,重度放牧区值最小,随后显著升高。9月,随牧压的升高,Pielou指数连续降低。10月均匀度指数表现先升高后降低的趋势。
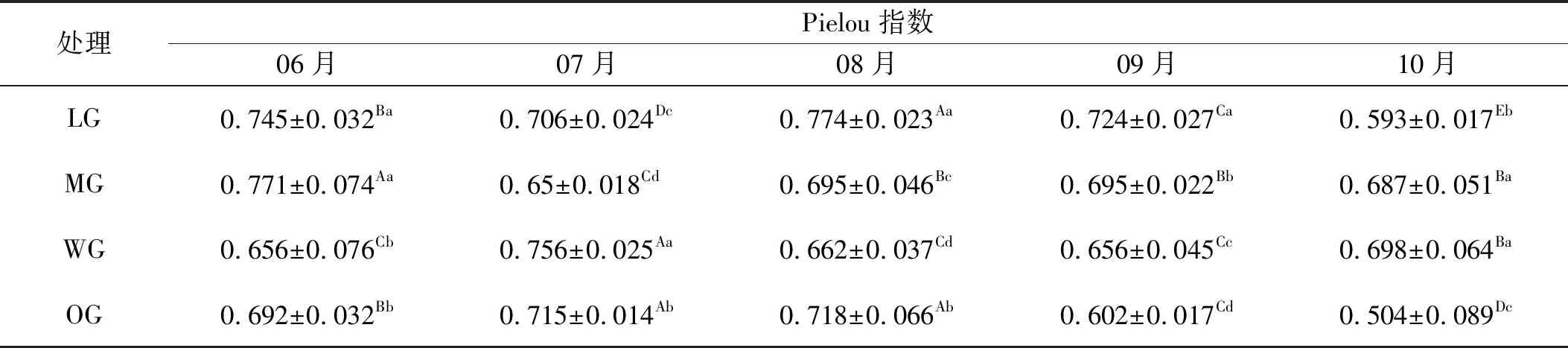
表5 有毒植物的Pielou指数
3 讨论
3.1 时间对有毒植物生物多样性的影响
研究不同放牧强度下有毒植物种群特征的变化,将为深入理解放牧等人类活动对草地生态造成的危害提供直接快捷的评价指标[15]。从时间尺度上有毒植物的丰富度表现出先降低再升高再降低的趋势。其中6月有毒植物丰富度较高,这可能与有毒植物生长速度快和草地基本情况有关。6月是植物返青期,此时有毒植物生长速度快于禾本科等牧草,在水肥资源竞争中占据优势地位。7~8月,可食牧草在与有毒植物的水肥竞争中逐渐取得优势,最终取代了有毒植物在群落中的优势地位。在9月植物枯黄期,对家畜而言食物资源的质量降低,选择性采食开始转变为被迫采食,所以有毒植物的丰富度降低。10月是牧草消亡期,有毒植物的重要值普遍低于其他月份。其原因可能是9月食物资源质量低,有毒植物和牧草都被采食殆尽,甚至在过度放牧区,植物的立枯体都被采食。在植物已经停止生长的10月份,植物无再生能力,所以无论是有毒植物还是牧草,其丰富度都大大降低。
研究表明植物有毒植物的生物多样性Shannon-Wiener指数和Peilou指数在时间尺度上表现出降低的趋势。这主要是由植物生长以及环境中水肥条件共同决定。在7月,水肥条件优越,植物进入高速生长期,有毒植物在6月积累的生长优势使其占据更多的水肥以及环境,同时,家畜采食引起的有毒植物超补偿生长机制使其在短时间内重建种群,所以在7月有毒植物生物多样性升高。8月植物生长进入盛草期,除重度放牧区外,其他试验区内有毒植物都达到了生物多样性的最高值。可食牧草的生长达到顶峰以及草地环境资源的累计和富余,会让毒草功能群为了巩固自己的生态位,产生一部分趁虚而入的季节性毒草。家畜的选择性采食达到顶峰,优良的可食牧草会被大量的选择性采食。这样,反而提高了有毒植物在整个群落中所占的比例。在枯黄期9月,草地生态系统植被基况出现巨大转变。大部分可食牧草营养物质显著降低,适口性变差。在牧草消亡期10月,尤其是在过度放牧区内,随着牧压的升高,一方面由于家畜的被迫采食,另一方面季节性毒草毒性的丧失和常年性毒草毒性的减弱,致使物种多样性严重降低。说明草地退化已经到不可控制的局面,有毒植物无法正常发挥维持生态系统结构和功能稳定性的作用[16]。
3.2 放牧对有毒植物多样性的影响
在轻度放牧区,有毒植物生物多样性指数较高。虽然伴随着水肥条件的改变,有毒植物的生物多样性指数存在一定波动,但均处于较高水平。其主要原因在于家畜的选择性采食。在轻度放牧区内,家畜会优先选择采食适口性好的牧草,为有毒植物快速在种群中占据优势地位提供生长空间,试验发现,黄花棘豆的丰富度较低。可能是因为黄花棘豆是豆科有毒植物,虽然有毒,但具有一定的营养价值[17]。轻度放牧区毒草有毒植物多样性指数均最大,说明适度干扰有毒植物对草地生产力和稳定性的维持发挥着重要作用[18]。但在过度放牧区的生物多样性指标均降低,表明逆境条件已经超出有毒植物的阈值,草地负荷严重超出正常水平,有毒植物已经开始表现出无法发挥维持生态系统功能与结构的稳定性的功能,这从侧面验证了毒草是草地植被逆行演替的产物。
在中度放牧区,有毒植物的物种丰富度,Shannon-Wiener指数和Peilou指数都保持在一个较低且稳定的水平上,说明有毒植物在群落中并未占据主要地位。适度的放牧践踏和采食,有利于可食牧草对水肥资源在草地群落中的竞争[19]。并且在采食牧草的过程中也会一定程度的采食有毒植物,为牧草提供生存空间,所以有毒植物的物种丰富度较低,符合“中度干扰假说”[20]。对比发现重度放牧牧区有毒植物的多样性在各时间尺度上都高于中度放牧区,这与李镇清[21]和李永宏[22]的研究结果一致。说明有毒植物对不同放牧强度的响应与牧草的响应是一致的。
3.3 有毒植物多样性与植物病害的关系
在过度和重度放牧区,草畜之间的动态平衡已经被打破,家畜过度的采食每种植物,是物种丰富度较低的主要原因。且由于极低的物种丰富度,不能对草地病害发生和流行设置障碍,致使过度和重度放牧区易发生病害[23-24]。由于食物资源的匮乏,家畜对食草地的利用方式由选择性采食变为被迫性采食,使每种植物上都有食痕;加之采食过程中无意识地对草地造成的践踏伤痕以及家畜便溺,使植物感染病毒以及真菌等病原体的概率大大增加[25],最终造成草地病害的发生与流行,进一步降低放牧区的有毒植物以及可食牧草多样性。相比重度放牧,过度放牧区中草畜平衡关系几乎完全被打破,感染病害的草也被采食,反而可以减轻某些病害。一些家畜易携带的病原微生物,却可以借助家畜在放牧区内游走的机会扩大传播面积[25-26]。在中度放牧区中,毒害草的物种丰富度、Shannon-Wiener指数和Peilou指数都较稳定。中度放牧区中适度的有毒植物丰富度以及家畜的中度干扰,可以降低病害发生的几率。一方面,对家畜采食已发生病害的牧草,减少初侵染源;另一方面,有毒植物对草地病害的传播设置天然屏障,阻隔牧草病害的传播,使其不能大面积流行。与此同时,中度放牧区内的采食强度,有助于缩短植物的生长期,减少了寄主植物被病原体侵染的机会[27]。
试验中所选取的家畜是高山细毛羊,所以其对草地利用方式与牛羊混牧的放牧区存在一定差异。高山细毛羊采食近地面的较低矮的植物,对植株采食更彻底,尤其是在轻度放牧区内,选择性采食达到顶峰,虽然家畜数目不多,但是依然可以对牧草形成高度的采食胁迫,为有毒植物生长提供最大的空间。研究结果表明,不同放牧区内生物多样性指数差异较大,但不同时间同一放牧区内差异较小,说明放牧是影响有毒植物生物多样性的重要因素。并且,高山细毛羊的体重远低于天祝县常见的白牦牛,同等牧压下,两种动物的践踏模式与践踏强度完全不同[20]。因此,白牦牛牧区有毒植物的群落特征可能与试验放牧区存在较大差异。如要继续研究祁连山东段地区有毒植物与放牧之间的关系,仍需要深入探讨不同放牧家畜在不同放牧强度下,对植被、土壤理化属性以及微生物等因素的影响。
4 结论
研究结果表明,不同放牧区内生物多样性指数差异较大,但不同时间内同一放牧区内差异较小,尤其是中度放牧区,有毒植物的3个生物多样性指数较低且趋于稳定,轻度放牧区和过度放牧区有毒植物生物多样性指数较高且变化较大,说明放牧是影响有毒植物生物多样性的重要因素,中度放牧强度更有利于草地的进展演替。试验中所选取的家畜是高山细毛羊,所以其对草地利用方式与牛羊混牧的放牧区存在一定差异。高山细毛羊采食近地面的较低矮的植物,对植株采食更彻底,尤其是在轻度放牧区内,选择性采食达到顶峰,虽然家畜数目不多,但是依然可以对牧草形成高度的采食胁迫,为有毒植物生长提供最大的空间。并且,高山细毛羊的体重远低于天祝县常见的白牦牛,同等牧压下,两种动物的践踏模式与践踏强度完全不同[21]。继续研究祁连山东段地区有毒植物与放牧之间的关系,仍需要深入探讨不同放牧家畜在不同放牧强度下,对植被、土壤理化性质以及微生物等因素的影响。
猜你喜欢
草食家畜(2024年1期)2024-02-06 09:59:00
今日农业(2021年12期)2021-10-14 07:30:52
今日农业(2021年10期)2021-07-28 06:28:00
今日农业(2020年14期)2020-12-14 19:47:34
电脑报(2020年12期)2020-06-30 19:56:42
——黔南扁穗雀麦
贵州农业科学(2019年1期)2019-02-20 02:09:54
癌症进展(2016年9期)2016-08-22 11:33:10
鹿鸣(2015年7期)2015-05-30 10:48:04
当代畜禽养殖业(2014年2期)2014-08-22 02:38:48
声屏世界(2014年6期)2014-02-28 15:18:07
