
祁连山东段不同类型灌丛斑块土壤特征对围封的响应
2019-11-18 06:17刘玉祯曹文侠王金兰李文辛雨琼王世林王小军
草业学报 2019年11期
刘玉祯,曹文侠,王金兰,李文,辛雨琼,王世林,王小军
(甘肃农业大学草业学院,草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
高寒灌丛作为高寒带的地带性植被广泛分布于青藏高原,多位于生境阴湿的高山带阴坡,其中以头花杜鹃(Rhododendroncapitatum)、山生柳(Salixoritrepha)和金露梅(Potentillafruticosa)为优势种的高寒灌丛草地最具代表性。高寒灌丛草地通常具有明显的层片结构,除浓密的灌木层外,还有较为发达的苔藓层和不发达的草本层。其特殊的生态序列位置使其拥有了较强的生态环境功能和较为丰富的生物多样性[1]。因此,高寒灌丛草地在青藏高原生态系统稳定中扮演着重要角色的同时,在能源替代、群落演替研究等方面也具有十分重要的意义[2]。土壤作为植被生长的载体,在诸多自然因素及人为因素的干扰下,其理化性质的空间分布常呈现出缀块型或梯度格局,也就导致了土壤理化性质在一定程度上具有随机性和相关性[3-5]。对于高寒灌丛草地而言,其土壤理化性质亦是如此,灌木的存在会导致土壤养分和水分通过地表径流与土壤侵蚀的作用逐渐汇集在灌下形成“肥岛”[6],从而形成了土壤理化性质在水平空间分布上的异质性。土壤理化性质不但是衡量土壤肥力和土壤质量的重要指标,也是调控植物生长发育的关键生态因子[7]。土壤化学性质,诸如有机碳、全效氮磷钾、速效氮磷钾等可直接影响土壤肥力,进而影响地上植被的生长,最终影响到高寒灌丛生态系统功能的发挥。因此,研究不同高寒灌丛斑块土壤理化性质的变化规律,可以了解土壤结构与功能,也能够明晰植被与土壤间相互协同的变化作用[8-9],这对于高寒灌丛草地生态过程研究等具有借鉴作用。
祁连山位于青藏高原东北缘,是我国西部重要的生态安全屏障[10],也是我国首批确定建设的国家公园。祁连山灌丛草地分布广泛,是当地牧民传统的冬春季放牧草场。但近年来,由于气候变化及过度放牧等因素[11-12],导致高寒灌丛草地出现不同程度的退化,进而影响着高寒灌丛草地生态系统的结构与功能[13],严重扼制了高寒灌丛草地生态系统的可持续发展。因此,亟待对祁连山退化高寒灌丛草地生态系统进行植被修复与生态重建。围栏封育作为最经济有效的恢复手段被广泛应用于祁连山退化高寒灌丛草地生态系统的修复中。因此,明晰典型高寒灌丛斑块土壤理化特征对围封的响应能够有效指导退化高寒灌丛草地生态系统的植被修复与生态重建。近年来,虽然已有学者对高寒灌丛进行相关研究,但是大多数研究均基于植物群落结构[14]、土壤微生物群落结构[15]、不同退化程度高寒灌丛草地土壤理化性质的变化[16]等。关于高寒灌丛斑块土壤理化性质及其分布特征对围封的响应报道较少。鉴于此,本研究以祁连山东段头花杜鹃、山生柳和金露梅3种高寒灌丛斑块土壤为研究对象,综合运用分析化学、统计学等研究手段,旨在探讨放牧利用后进行围封恢复会对高寒灌丛斑块土壤理化性质分布规律产生何种影响,从而揭示不同类型高寒灌丛斑块土壤理化性质变化规律对围封的响应,以期为祁连山退化高寒灌丛生态系统植被恢复与重建、管理及可持续利用等提供基础数据和科学理论依据。
1 材料与方法
1.1 研究区自然概况
研究区位于祁连山东段(37°10′14.36′′ N,102°47′12.13′′ E),平均海拔在2960 m以上。气候类型隶属于大陆性高原寒温带季风气候,年均降水量为414.98 mm,降水多为地形雨,66%的降水集中在7-9月。全年无绝对无霜期,仅分为冷热2季,年均温为0.13 ℃,最冷月平均气温-11.4 ℃,最热月平均气温11.2 ℃[17]。土壤类型隶属于高山灌丛草甸土且土层较薄(40~80 cm)。植被沿海拔方向分异特征明显,依次分布的典型灌木有金露梅、山生柳、头花杜鹃等;依次分布的典型草本植物有珠芽蓼(Polygonumviviparum)、问荆(Equisetumarvense)、苔草(Carexspp.)、嵩草(Kobresiaspp.)等。
1.2 研究方法
1.2.1样地设置及样品采集 于2014年6月选取坡向、海拔等基况基本一致的冬春季放牧高寒灌丛草地作为研究样地,并沿海拔方向设置3个100 m×30 m的围栏,各围栏间距均在40 m以上。以围栏外的放牧地作为对照,围栏内为禁牧样地,全年禁牧;围栏外为放牧样地,全年放牧率约在5.8~6.7羊单位·hm-2。放牧样地放牧家畜为当地牦牛和藏羊。围栏内外灌下植被基本概况见表1。
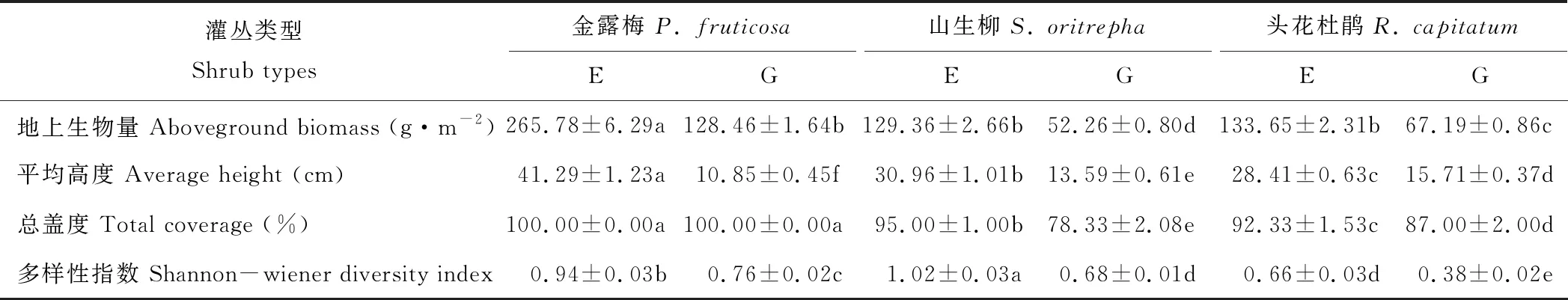
表1 围栏内外灌下植被基本概况Table 1 General information of undergrowth of shrubs inside and outside fences
注:E,禁牧样地;G,放牧样地。同行不同小写字母表示在5%水平上差异显著(P<0.05),下同。
Note: E, exclosure plot; G, grazing plot. Different small letters in the same column show significant differences (P<0.05), the same below.
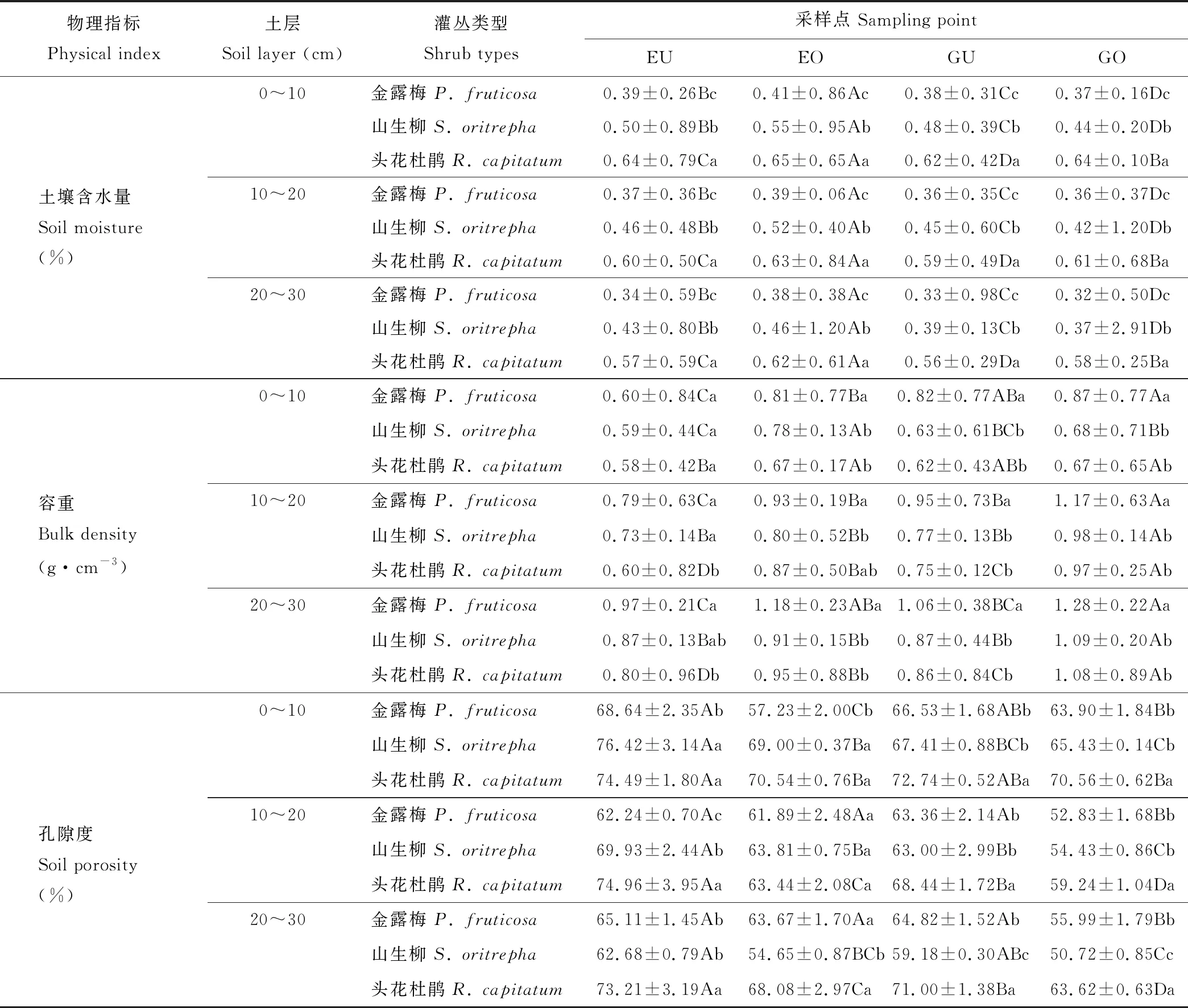
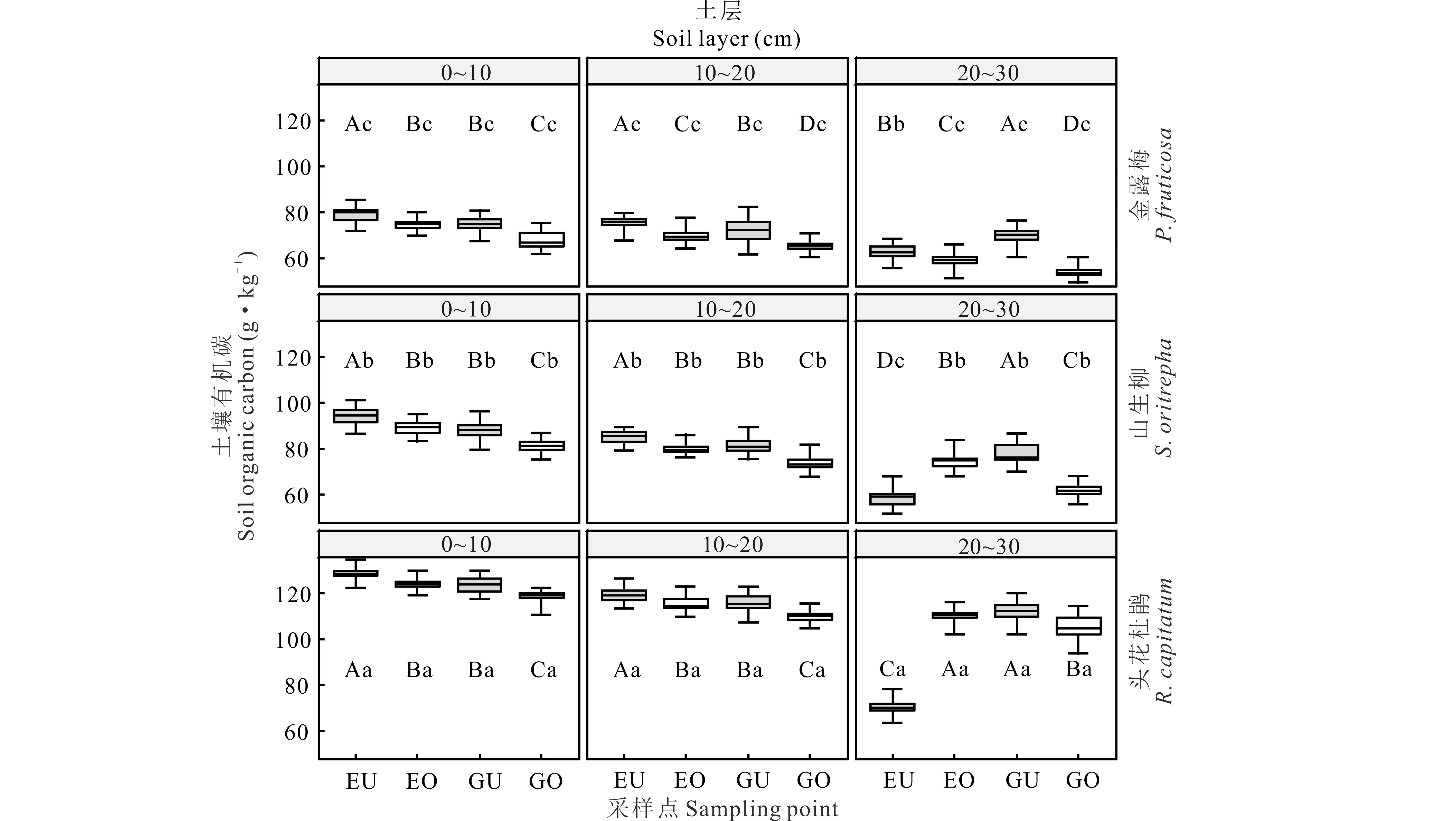
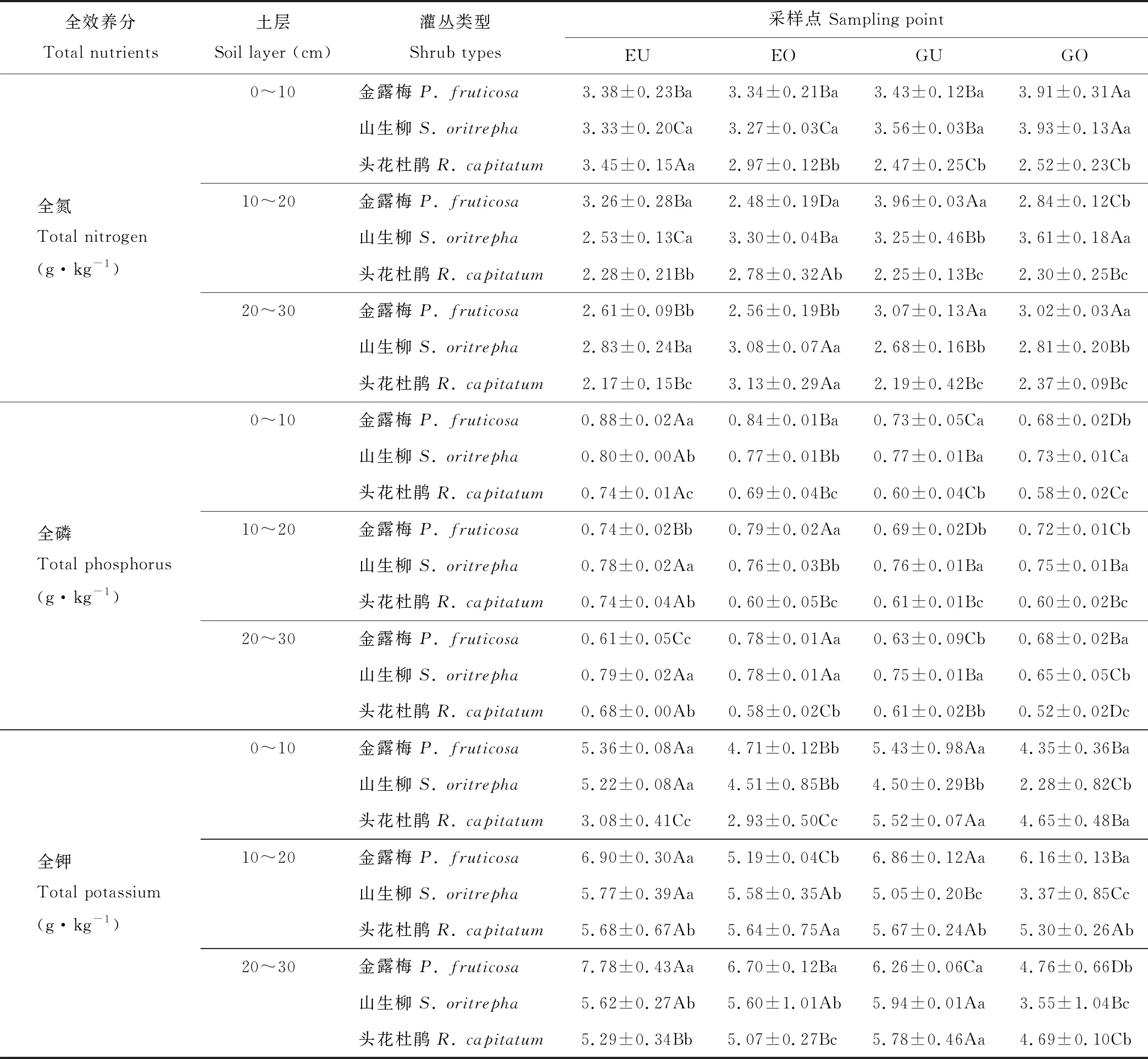
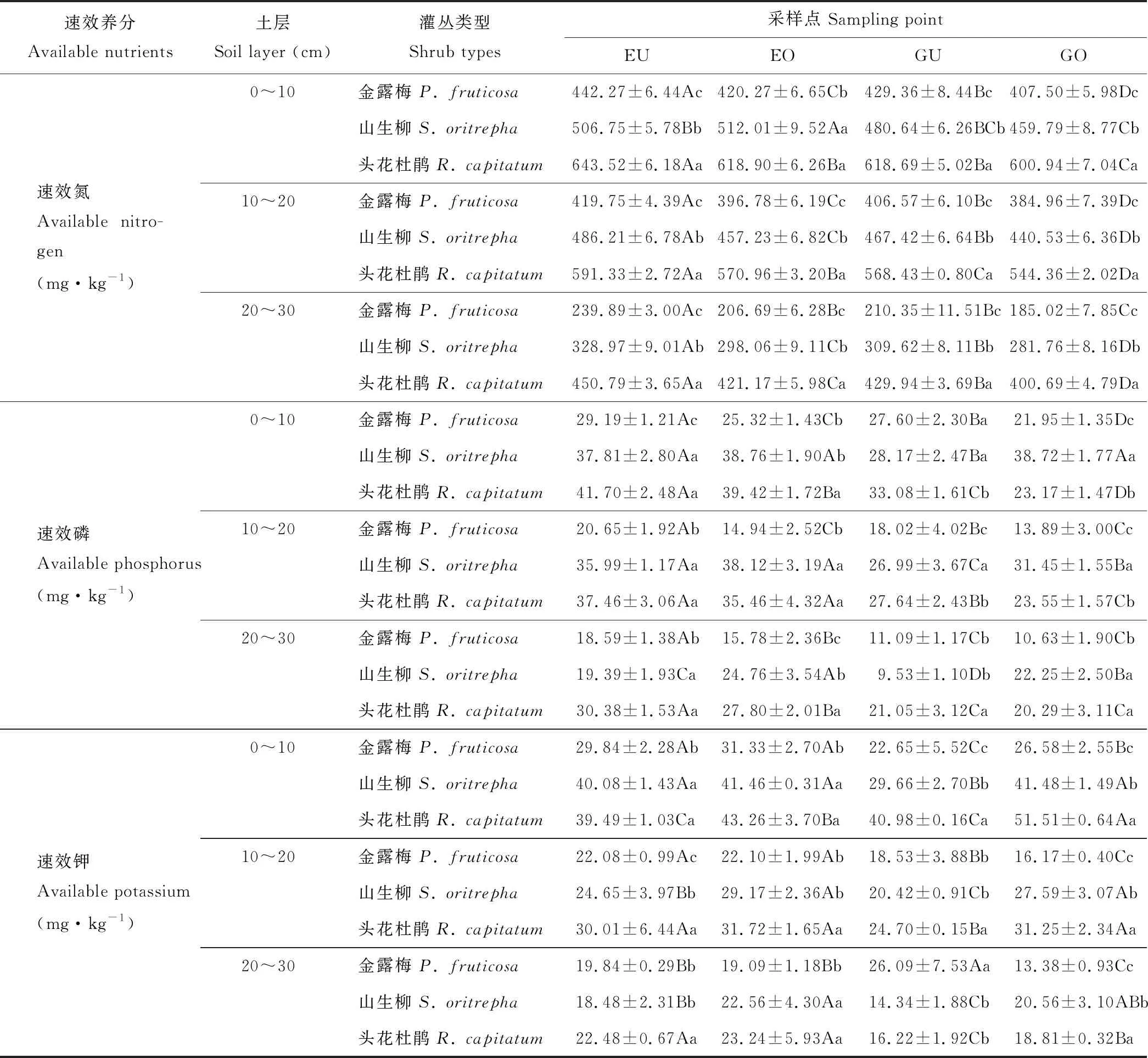
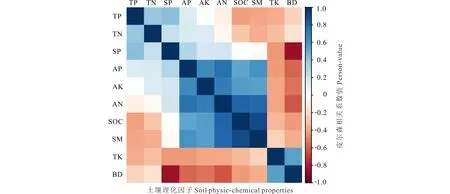
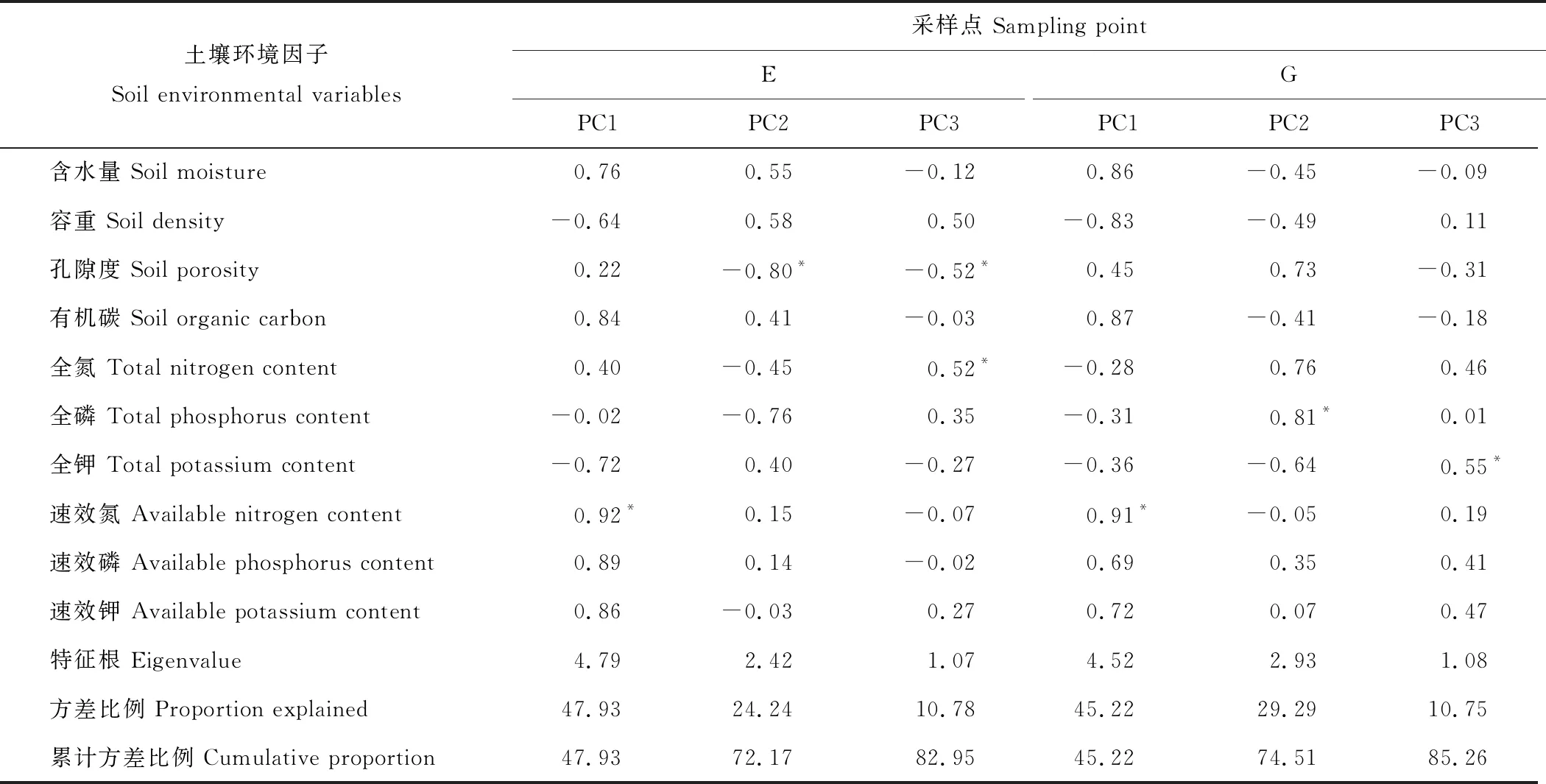
土壤样品采集工作于2018年7月15日进行,采集前先进行植被调查并记录不同灌丛斑块的高度、冠幅和冠基,以预先的调查结果为基础,在各围栏内外分别选取中型金露梅灌丛斑块(14 cm3 (1) 式中:H代表灌丛高度;D代表灌丛冠幅平均直径[18]。采用灌丛斑块个体体积V的立方根V′来界定灌丛体型大小。 1.2.2样品分析 采用环刀法测定土壤含水量(soil moisture, SM),采用烘干法测定容重(bulk density, BD);通过土壤容重与土壤比重来计算土壤孔隙度(soil porosity, SP)[19];土壤有机碳(soil organic carbon, SOC)的测定在有机碳分析仪(multi N/C 2100s,Analytik Jena Germany)中进行;参照鲍士旦[20]的方法:采用凯氏定氮法测定全氮(total nitrogen, TN);采用钼锑抗比色法测定全磷(total phosphorus, TP);采用氢氧化钠熔融-火焰光度计法测定全钾(total potassium, TK);采用碱解氮扩散法测定速效氮(available nitrogen, AN);采用碳酸钠浸提-钼锑抗比色法测定速效磷(available phosphorus, AP);采用乙酸铵浸提-火焰光度法测定速效钾(available potassium, AK)。所有土壤样品指标的测定均重复3次求取平均值作为研究的原始数据。 首先,运用Shapiro-test函数对原始数据进行正态分布检验,若有不满足正态分布的数据则对其进行对数转换。随后,运用One-way ANOVA分析对围栏内外同一类型灌丛斑块(U点、O点)、不同类型灌丛斑块同一土层的土壤理化性质进行差异性检验,再运用Tukey-HSD方法进行均值的多重比较。鉴于本研究涉及到10个变量,过多的变量将不利于合理的分析和解释问题。因此,本研究选择运用主成分分析(principal component analysis, PCA)进行降维处理。基于提取轴信息的判别标准如下[21]:1)当排序轴获得绝大部分解释方差时,即累积解释方差比例超过66%(图4);2)采用Kaiser-Guttman标准,即选取特征根大于1的轴提取排序轴(表5)。采用Person相关性进行各土壤理化性质间的相关性分析,相关性大小采用Student’s T 检验。数据统计分析及绘图均在R 3.6.0(R Development Core Team,2016)中进行。 图1 采样点示意图Fig.1 Schematic diagram of sampling points U1、U2、U3、U4:灌下采样点 Sampling point under the shrubs;O1、O2、O3、O4:灌外采样点 Sampling point outside the shrubs canopy; EU:围栏内灌下采样点 Sampling point under the shrubs inside the exclosure;EO:围栏内灌外采样点 Sampling point outside the shrubs canopy inside the exclosure;GU:围栏外灌下采样点 Sampling point under the shrubs inside the grazing plot;GO:围栏外灌外采样点 Sampling point outside the shrubs canopy inside the grazing. 3种灌丛斑块土壤物理性状在0~10 cm、10~20 cm、20~30 cm土层中及EU、EO、GU、GO点均有显著的变化趋势(表2)。其中,土壤含水量整体上表现为EU 表2 围栏内外不同类型高寒灌丛斑块土壤物理性质空间分布Table 2 Spatial distribution of soil physical properties around different alpine shrubs patch with enclosure and grazing 注:EU,围栏内灌下采样点;EO,围栏内灌外采样点;GU,围栏外灌下采样点;GO,围栏外灌外采样点。同行不同大写字母表示在5%水平上差异显著(P<0.05),不同灌丛斑块类型同一土层标有不同小写字母表示在5%水平上差异显著(P<0.05),下同。 Note: EU, sampling point under the shrubs inside the exclosure; EO, sampling point outside the shrubs canopy inside the exclosure; GU, sampling point under the shrubs inside the grazing plot; GO, sampling point outside the shrubs canopy inside the grazing. Different capital letters in the same column show significant differences (P<0.05), different small letters in the same soil layers show the significant differences (P<0.05), the same below. 从不同高寒灌丛斑块土壤有机碳含量变化趋势可以看出(图2),在20~30 cm土层中,有机碳含量变化在各灌丛斑块中均呈现出EU 3种灌丛斑块土壤全N含量变化规律在0~10 cm土层较为一致(表3),除头花杜鹃灌丛斑块为EU>GU、EO>GO外,均为EU>EO、GU 图2 围栏内外不同类型高寒灌丛斑块土壤有机碳空间分布Fig.2 Spatial distribution of soil organic carbon around different alpine shrubs patch with enclosure and grazing 同类型灌丛斑块不同大写字母表示在5%水平上差异显著(P<0.05),不同灌丛斑块类型同一土层标有不同小写字母表示在5%水平上差异显著(P<0.05),下同。Different capital letters in the same shrubs patch show significant differences (P<0.05), different small letters in the same soil layers show the significant differences (P<0.05), the same below. 对比灌下与灌外,3种灌丛斑块土壤速效N和速效P含量在0~10 cm、10~20 cm、20~30 cm土层中大体上表现为EU>EO、GU>GO,速效K含量则大体上表现为EU 虽然3种不同类型的灌丛斑块在围栏内外及灌下、灌外土壤理化性质的相关性不尽相同,但是总体上依旧存在一些较为一致的变化规律(图3):围栏内外及灌下、灌外土壤容重与孔隙度呈显著负相关,土壤有机碳与土壤含水量及速效氮呈显著正相关,与容重呈负相关,土壤含水量与全效养分呈显著负相关,但与速效养分却呈显著正相关。 表3 围栏内外不同类型高寒灌丛斑块土壤全效养分空间分布Table 3 Spatial distribution of soil total nutrients around different alpine shrubs patch with enclosure and grazing 表4 围栏内外不同类型高寒灌丛斑块土壤速效养分空间分布Table 4 Spatial distribution of soil available nutrients around different alpine shrubs patch with enclosure and grazing 图3 围栏内外不同类型高寒灌丛斑块土壤理化性质相关性分析Fig.3 Correlation analysis of the soil physic-chemical properties around different alpine shrubs patch with enclosure and grazing TP:全磷 Total phosphorus;TN:全氮 Total nitrogen;SP:土壤孔隙度 Soil porosity;AP:速效磷 Available phosphorus;AK:速效钾 Available potassium;AN:速效氮 Available nitrogen;SOC:有机碳 Soil organic carbon;SM:土壤含水量 Soil moisture;TK:全钾 Total potassium;BD:容重 Bulk density.下同 The same below. 由主成分分析可知(图4,表5),围栏内外排序轴前3轴累计方差解释比例分别为82.95%和85.26%,且特征根均大于1,表明土壤理化性质在前3排序轴的载荷值较高。本研究提取解释率相对较高的前2轴为下一步分析做准备。围栏内第一主成分累积贡献率为47.93%,主要体现了速效氮(0.92)的贡献,第二主成分累积贡献率为24.24%,主要体现了孔隙度(-0.80)的贡献;围栏外第一主成分累积贡献率为45.22%,主要体现了速效氮(0.91)的贡献,第二主成分累积贡献率为29.29%,主要体现了全磷(0.81)的贡献。由图4可以看出,头花杜鹃灌丛斑块在围栏内外始终保持着较好的土壤理化性质。 图4 围栏内外不同类型高寒灌丛斑块土壤理化性质的PCA分析Fig.4 Principal component analysis of the soil physic-chemical properties around different alpine shrubs patch with enclosure and grazing a:禁牧样地 Exclosure plot;b:放牧样地 Grazing plot. 土壤理化性质作为维系高寒灌丛生态系统群落稳定的重要因素之一,亦是调控植被生长发育的关键生态因子[22]。本研究表明,同类型灌丛斑块就物理性状而言,总体表现为灌下显著优于灌外,围栏内又显著优于围栏外(除含水量在围栏内表现为灌下显著低于灌外)。围栏内外头花杜鹃灌丛斑块均有着最高的含水量、孔隙度和最小的容重,表明相对于其他两种灌丛斑块,头花杜鹃灌丛斑块拥有更好的物理性状,与前人研究结果一致[23]。相较于围栏外的放牧地,围栏内由于没有放牧干扰,草本植物拥有良好的生长条件,而灌下草本植物由于灌木的遮阴等作用致使其盖度和生物量不及灌外草本,因此,灌外草本的覆盖度更高、蒸发量更小,从而提高了灌外土壤的蓄水能力,导致围栏内灌外土壤含水量要高于灌下土壤含水量。围栏外则由于灌木冠幅的庇护作用,使得家畜采食时多以灌外草本为主,从而致使灌下草本盖度及生物量高于灌外草本,且由于家畜的采食和践踏导致灌外土壤植被盖度低的同时土壤紧实度也较高,发生降水时更容易形成地表径流,致使灌外的蓄水能力降低。因故,围栏外灌下土壤水分含量高于灌外。不同类型灌丛斑块土壤水分含量差异形成的原因与其分布位置密切相关,相较而言,头花杜鹃灌丛分布海拔最高,生境本就阴冷潮湿,外加郁闭度较高,家畜的采食扰动也较小,其阴湿环境为苔藓地被物等提供了良好的积蓄条件,大量地被物的存在可有效提高其蓄水能力,使头花杜鹃灌丛斑块拥有了较高的持水能力、较小的容重及较大的孔隙度。金露梅灌丛斑块分布海拔较低,受外界干扰程度最为强烈,所以其物理性状最差;山生柳灌丛斑块分布海拔通常介于二者之间,受干扰程度次之,从而造就了头花杜鹃灌丛土壤物理性质最优的分布特征。 表5 主成分分析(PCA)中前3轴环境变量的载荷和解释方差Table 5 The loadings and explained variance of environmental variables in the first three axes in PCA 注:*表示该变量的载荷绝对值在排序轴中排序第一。 Note:* indicates that the absolute value of the load of this variable ranks first on the ordering axis. 诸多研究已经证明灌丛“肥岛”现象广泛存在于干旱半干旱区草地生态系统中[24-25]。本研究通过分析不同类型灌丛斑块围栏内外灌下和灌外土壤有机碳及土壤养分的差异发现,分布于祁连山的3种典型高寒灌丛斑块亦是如此。土壤有机碳含量是评定土壤肥力高低的重要指标,主要来源于凋落物、动物残体及废弃物的归还等[26],同时也会在一定程度上受到微生物活动的影响。研究区不同灌丛斑块有机碳含量总体上表现为头花杜鹃灌丛斑块>山生柳灌丛斑块>金露梅灌丛斑块,且灌下显著优于灌外,围栏内又显著优于围栏外。这是由于围栏内无放牧干扰,因此,相较于围栏外其灌下、灌外均有较多的凋落物返还,也就导致围栏内有机碳含量显著高于围栏外。围栏内灌下有机碳含量显著高于灌外,这是由于灌下不仅有灌木的枯枝落叶输入土壤,还有草本植物凋落物的归还及一部分鸟类栖息时的排泄物归还等,从而造就了围栏内灌下有机质含量显著高于灌外的分布特征。围栏外灌下有机质含量显著高于灌外则主要是由于放牧家畜采食导致,家畜的采食作用导致灌下凋落物输入高于灌外,即便灌外拥有着动物排泄物的归还,但是由于其土壤物理性质较差,有机质易随降水时形成的地表径流流失,从而导致围栏外灌下有机质含量依旧高于灌外。不同类型灌丛斑块有机质含量的差异与其分布生境依旧密切相关,相较于山生柳灌丛斑块和金露梅灌丛斑块,头花杜鹃灌丛斑块的地表植被分布状况最差,但是由于其分布生境阴冷潮湿的原因,一方面致使其有着大量地被物的存在,另一方面,低温抑制了微生物的活动,导致积蓄在地表的动植物残体等分解缓慢,逐渐转化积累,从而造就了头花杜鹃灌丛斑块有机碳含量最高的分布特征。 李冬林等[27]研究表明,土壤氮素含量的高低一定程度上取决于有机质的含量,而有机质的60%~80%又是由有机碳构成的,因此,有机碳含量较为丰富的灌丛斑块氮素含量也相对较高。本研究表明,有机碳含量变化对全氮含量的影响不明显,但是有机碳含量较高的灌丛斑块速效氮含量也较高,即围栏内外不同类型灌丛斑块速效氮含量与土壤有机碳分布格局基本一致。 土壤磷素作为植被生长发育的必需元素之一,自然状态下其消长主要通过磷的矿化、生物迁出及土壤淋融作用[28]。土壤磷素一般在土壤发育时间较长或高度淋融的地区较为缺乏,并且会成为植被生长的限制性因子。本研究中,土壤全磷总体上在围栏内外均表现为0~10 cm土层灌下大于灌外,10~20 cm、20~30 cm土层灌下小于灌外。分析认为,表层土壤较高的磷含量为灌下草本植被提供了良好的生长条件,同时,为了避免与草本植物表层磷素的竞争,灌木则会趋向于将根系延伸至更深层的土壤,从而导致了全磷在不同土层的分布差异,也说明了高寒灌木一般都拥有着较大的地下生物量。土壤速效磷变化规律与有机碳分布格局基本一致,主要表现为灌下显著高于灌外、围栏内显著高于围栏外。Walbridge[29]的研究表明,土壤微生物可将植物难以吸收利用的磷通过生化作用转化为较易吸收利用的形态,而灌丛下良好的微生境为微生物的活跃提供了重要的环境,因此灌下磷素含量显著高于灌外。土壤速效磷之所以会呈现出头花杜鹃灌丛斑块>山生柳灌丛斑块>金露梅灌丛斑块的分布规律,依旧与其分布位置密切相关。虽然头花杜鹃灌丛斑块的低温抑制了微生物的活动,但是由于其受干扰程度最小且土壤物理性状最佳,致使其灌丛斑块中土壤微生物通过生化作用转化成的土壤速效磷不易流失,而山生柳灌丛斑块和金露梅灌丛斑块微生物活动虽然比较活跃,但是较为强烈的干扰和较差的物理性状更易导致土壤速效磷的流失。 土壤钾素是植被进行光合作用的重要元素之一,植被主要吸收利用的是速效钾,但是速效钾只占土壤全钾的0.1%~2.0%[30]。土壤中的全钾主要来源于土壤母质成土过程中的释放,拥有较强的可溶性且极易流失[31]。本研究中,各灌丛斑块土壤全钾含量总体上呈现出围栏内高于围栏外、灌下高于灌外的特征。围栏内土壤物理性状显著优于围栏外,因此,围栏外的钾素更易流失,从而导致围栏内土壤全钾含量显著高于围栏外;灌下土壤全钾含量高于灌外的原因亦是如此。各灌丛斑块土壤速效钾含量总体上呈现出围栏内高于围栏外、灌下低于灌外的特征。土壤速效钾之所以会产生围栏内显著高于围栏外的分布特征与其极易流失的特征密切相关,而灌下显著低于灌外则是由于灌下土壤速效钾由灌木以及灌下草本共同吸收利用,从而造就了灌下低于灌外的分布特征。土壤速效钾之所以会呈现出头花杜鹃灌丛斑块>山生柳灌丛斑块>金露梅灌丛斑块的分布规律,仍旧与其分布位置密切相关。相关研究表明,植被类型与干扰强度是调控土壤中速效钾含量的主要因子[32]。头花杜鹃灌丛分布海拔较高,山生柳灌丛次之,金露梅灌丛则最低,导致其受干扰程度不同,与此同时,3种灌丛斑块中分布着不同类型的植被。因此,在局部生境差异及外界干扰程度差异的共同作用下影响了土壤速效钾在不同灌丛斑块间的再分配。 土壤理化性质在一定程度上具有相关性。通过相关性分析发现,虽然3种不同类型的灌丛斑块在围栏内外及灌下、灌外的土壤理化性质相关性各不相同,但是依旧存在一些较为一致的变化规律。不同水平下土壤容重与孔隙度均呈显著负相关,与前人研究结果一致[15,33]。另外,土壤有机碳与土壤含水量和多个土壤化学性质呈显著正相关,与容重呈负相关。土壤有机碳可有效改善土壤物理结构,增加土壤的通透性,因故,土壤有机碳含量越高土壤容重就越低。相关研究表明,土壤中氮素和钾素主要来源于有机质的矿化和分解,因此,有机质可以显著调控土壤中养分含量的变化[34]。土壤中的水分循环可直接影响植被的生长及土壤中各物质的转化过程[30]。本研究中,土壤含水量与全效养分呈显著负相关,与速效养分呈显著正相关,说明较高的含水量利于土壤氮磷钾的质量分解。 主成分分析致力于定量数据的分析,通过展示出原始数据的欧氏距离及线性关系而达到降维的目的。其核心在于降维后突出综合指标的影响且信息之间不重复,弱化变量之间自相关引起的系统误差,最终达到对研究中土壤理化性质精确分析的目的[35]。本研究发现,速效氮在围栏内外均是影响3种灌丛斑块及其植被分布的关键环境因子。未受放牧干扰时,影响祁连山东段3种灌丛斑块及其植被分布的重要环境因子主要为速效氮与孔隙度;受放牧干扰后则变为速效氮与全磷。头花杜鹃灌丛斑块在围栏内外始终保持着较好的理化性质,受放牧干扰后,头花杜鹃灌丛斑块样方排序距离与山生柳灌丛斑块和金露梅灌丛斑块的排序距离远离,表明头花杜鹃灌丛斑块拥有良好的抗外界干扰能力。 研究结果表明,祁连山东段3种灌丛斑块10个土壤理化性质中5个指标在灌下、灌外差异显著(土壤含水量、有机碳、全磷、全钾、速效氮)、6个指标在围栏内外差异显著(土壤含水量、容重、有机碳、全磷、速效氮、速效磷)。围栏内外土壤含水量与全效养分呈显著负相关,但与速效养分却呈显著正相关。主成分分析进一步表明,不论是否受到放牧干扰,速效氮均是决定祁连山东段3种灌丛斑块及其植被分布的关键环境因子;放牧干扰移除后,速效氮与孔隙度成为决定祁连山东段3种灌丛斑块及其植被分布的重要环境因子;受放牧干扰后,速效氮与全磷则成为决定祁连山东段3种灌丛斑块及其植被分布的重要环境因子。头花杜鹃灌丛斑块抗外界干扰能力较强,不论是否收到放牧干扰均能保持较好的土壤理化性质。短期围封能够显著增加灌丛斑块土壤含水量和速效养分含量,从而改善立地条件,也进一步证明围封更加有利于退化灌丛草地的恢复与重建。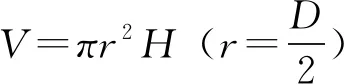
1.3 数据处理及分析

2 结果与分析
2.1 围栏内外不同类型高寒灌丛斑块土壤物理性状变化
2.2 围栏内外不同类型高寒灌丛斑块土壤有机碳含量变化
2.3 围栏内外不同类型高寒灌丛斑块土壤全效养分含量变化
2.4 围栏内外不同类型高寒灌丛斑块土壤速效养分含量变化
2.5 围栏内外不同类型高寒灌丛斑块土壤理化性质相关性分析
2.6 围栏内外不同类型高寒灌丛斑块土壤理化性质的PCA分析

3 讨论
3.1 不同水平土壤物理性质对比分析
3.2 不同水平土壤化学性质对比分析
3.3 土壤理化性质的相关性与PCA分析
4 结论
猜你喜欢
水土保持学报(2022年5期)2022-10-10
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
今日农业(2021年2期)2021-11-27
心声歌刊(2021年3期)2021-08-05
红蜻蜓·低年级(2020年12期)2020-06-20
小聪仔(幼儿版)(2020年12期)2020-02-01
好孩子画报(2019年8期)2019-09-19
民族音乐(2018年5期)2018-11-17
乡村地理(2018年3期)2018-11-06
发明与创新·大科技(2018年2期)2018-03-17
