
不同大花类群淫羊藿属植物的花部特征及生殖特征研究
2019-11-15 08:17陈兰英
植物研究 2019年6期
陈兰英 肖 肖 肖 娟*
(1.西华师范大学生命科学学院,南充 637000; 2.西华师范大学环境科学与工程学院,南充 637000)
花作为被子植物的繁殖器官具有很强的变异性,通过花性状能影响植物的传粉方式和交配模式[1]。其中,花部综合特征是影响传粉者有效传粉的本质因素,它可能会增加或者减少花粉的可授性,最终直接影响到结实率[2~3]。传粉是植物有性生殖过程的基础,异交植物通过花部特征中的花色、大小、花粉或者花蜜的报酬等来吸引昆虫进行传粉,花部特征的改变会引起传粉者种类和传粉行为的改变。目前,对于植物开花物候和繁殖特性方面的研究已有较多的报道,主要集中在同一物种不同微环境下的植物开花物候、结果特性及传粉繁育系统方面的研究[4~7],而对不同区域范围内同一种属的植物进行大尺度繁殖特性的研究内容鲜有报道。
淫羊藿为小檗科(Berberdiaceae)淫羊藿属(Epimedium)多年生宿根草本植物,该属植物主要分为大花类群和小花类群两大类,作为一种传统中药使用历史悠久[8~9]。其耐寒性极好,春日开花,花朵色彩美丽,形状多样,亦可用来做地被观赏植物,国外已培育出诸多庭院观赏品种[10~11]。该属植物虽以无性繁殖为主,但有性繁殖是无性繁殖的前提。研究表明,长期无性繁殖会影响植物的遗传多样性,遗传同一性会使后代个体更容易受到病虫害攻击;同时人为滥采滥挖行为加剧了淫羊藿野生资源的匮乏,其生物多样性受到了严重破坏[12~13]。有性繁殖能维持植物较高的遗传多样性,研究不同区域内同一属种植物的繁殖特性,通过人为种植来减少野生淫羊藿的采挖,保护野生资源非常有必要。国外学者对该属植物的传粉系统已有深入研究,但不同生态环境下,该属植物的生物学特性和传粉昆虫资源不尽相同,我国虽是淫羊藿属植物的分布中心,但不同属种淫羊藿之间传粉生态学对比研究的资料较少[14]。本研究以野生巫山淫羊藿、粗毛淫羊藿和宝兴淫羊藿为材料,探讨了三者的花部特征、访问者与生殖特性,旨在探讨不同淫羊藿属植物的生殖空间差异性,花部特征和传粉者对生殖成功的影响,为该属植物种间杂交培育新品种和物种遗传资源保护提供理论依据。
1 材料与方法
1.1 试验材料与研究地概况
巫山淫羊藿(E.wushanense)研究地位于四川省南充市高坪区、蓬安县及广安市岳池县之间的嘉陵江流域金城山国家森林公园,该地区冬季温和、夏季炎热多雨,属典型的季风湿润气候类型[15]。海拔560~900 m,年均气温17.6℃,全年降雨量约1 063 mm,1月均温5.4℃,7月均温21.7℃,植物生长期较长。
粗毛淫羊藿(E.acuminatum)位于四川省峨眉山市万年寺附近,该区域气候温暖,云雾多,日照少,降雨丰富,海拔1 000 m左右,属于暖湿气候带,一月平均气温约6.9℃,七月平均气温26.1℃。
宝兴淫羊藿(E.davidii)为四川省特有种,目前仅在四川省的宝兴县、峨嵋山市、天全县、和理县等地有分布。研究地位于四川省宝兴县蜂桶寨自然保护区,海拔1 000~4 000 m,全年均温5.9~7.2℃,无霜期228~242 d,年均降水量介于700~1 300 mm,降水多集中于夏季。
在以上3个研究地内选择数量较多的淫羊藿属植物种群为试验材料。
1.2 样地设置与生境因子测量
在每个研究地内分别设置3个样地(20 m2×20 m2),共计9个样地进行生境因子测量,测量内容包括:海拔、温度、空气相对湿度、土壤含水量、土壤有机质含量、相对光照强度等。温度、空气相对湿度利用法国KIMO温度湿度计测量;土壤含水量采用便携式土壤水分速测仪(TRIME-TD RZ)测定;土壤有机质含量采用硫酸重铬酸钾滴定法测定;相对光照强度(林下光照强度/裸地光照强度×100%)利用光照计(美国3413F-Field Scout)测量。
1.3 开花物候与花部特征观察
从群体、个体、花序和单花4个水平分别对3种淫羊藿的开花物候进行观察。具体方法:在淫羊藿花朵未开之前,每个种群选取30株,每株选一个花序,分别用不同颜色的线进行标记,且每株用标记牌进行编号。每隔2 d对同一花序的花蕾总数、隔天开放花朵数、剩余花蕾数,单花花期进行统计记录。在每一种群随机选取30株开花植株,对3种淫羊藿属植物的株形、花和叶进行性状观察并摄像,利用游标卡尺测量花部特征。
1.4 观察访花者
在3种淫羊藿属植物盛花期期间,各选取观测3个样方(2 m×2 m)内的开花植株进行标记,确定主要传粉昆虫并定点观察访花昆虫。8:00~18:00期间,跟踪观察并记录访花昆虫种类、访问次数,单次访花时间、访花行为的活动规律及天气变化。观察完成后,用昆虫网捕获访花昆虫标本带回实验室鉴定。
1.5 花部特征诱物试验
在3种淫羊藿的盛花期,对尚未盛开的花蕾套袋,待花朵即将盛开而雄蕊未裂时,作去距处理,每次观察访花前将袋子揭开观察并记录访花昆虫种类、访问次数,单次访花时间、访花行为的活动规律,之至结实后统计结实率。
1.6 柱头可授性及繁殖系统研究
在研究地连续两年进行了人工授粉试验,根据Dafni[16]的方法设立了如下10个试验组:①自然对照,不作任何处理,挂牌直至果实成熟;②套袋对照,不作处理,检验是否自花授粉;③人工自花授粉后,去雄,套袋,鉴定是否自交亲和;④同种同株人工异花授粉;⑤去雄不套袋,判断是否虫媒异交及酬物;⑥开花1~5 d同种异株人工异花授粉。
1.7 数据分析
运用SPSS23.0进行多因素方差分析、双变量相关性分析,按不同的植物种类进行LSD多重比较。
2 结果与分析
2.1 生境因子
3种淫羊藿属植物的生境因子,除相对光强和相对湿度无显著差异外,其他环境因子差异显著。3种植物材料的海拔、温度差异显著(P<0.05),其中,宝兴淫羊藿的海拔最高,其次是粗毛淫羊藿,巫山淫羊藿最低;温度为巫山淫羊藿>粗毛淫羊藿>宝兴淫羊藿。粗毛淫羊藿的土壤含水量和土壤有机质含量(P<0.05)均显著高于巫山淫羊藿及宝兴淫羊藿,而巫山淫羊藿与宝兴淫羊藿之间无显著差异(P>0.05)。
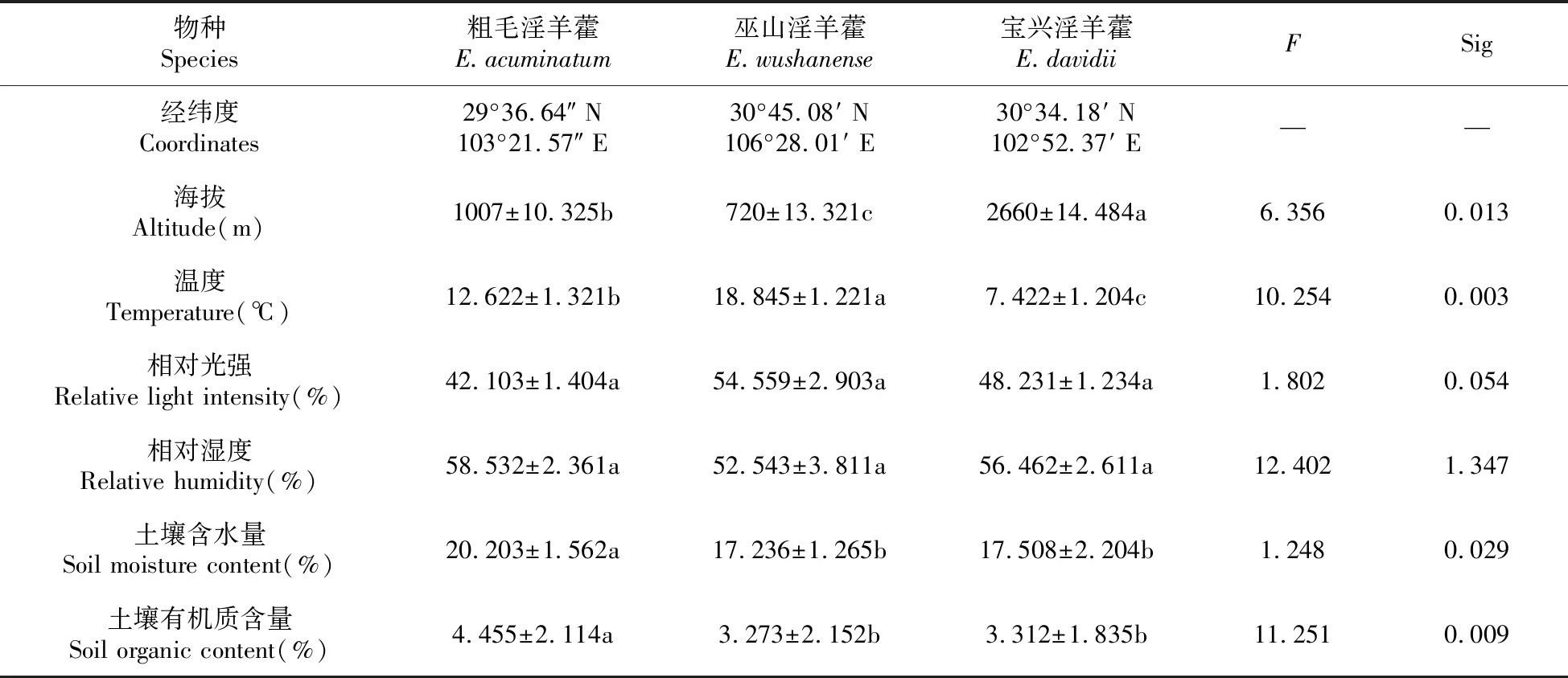
表1 不同淫羊藿属植物生境因子概况
注:表中数据为平均值±标准差(n=3),同行中不同字母表示处理间差异显著(P=0.05)。
Note:Values are means±SD(n=3),different letters in the same row mean significant differences among treatments atP=0.05 level.

图1 3种淫羊藿单花开花过程Fig.1 The flowering-process of single flower in three Epimedium species

图2 3种淫羊藿的有效访花者 A~B.巫山淫羊藿的访花者熊蜂属1和熊蜂属2;C.粗毛淫羊藿访花者熊蜂属3;D.宝兴淫羊藿隧蜂;E.拟蜂眼蝇;F.食蚜蝇Fig.2 Effective pollinators of of three Epimedium A~B.Bumblebees of Bombus sp1. and Bombus sp2. of E.wushanense; C.Bumblebees of Bombus sp3 of E.acuminatum; D.Argalictus resurgens; E.Cerioides annulata; F.Syrphidae of E.davidii
2.2 花部特征与单花开花过程
3种试验材料具有性和无性生殖能力。花为两性花,无特殊香味,倒立生长,距上有蜜腺,花朵部分器官(花序轴、单花花柄、内萼片、花距、花药及胚珠)在形态和数量上差异明显(图1)。花序轴和单花花柄中宝兴淫羊藿和巫山淫羊藿为密被毛;粗毛淫羊藿为疏生毛。宝兴淫羊藿的内萼片披针形,基部紫色,颜色由浅紫变为紫色;粗毛淫羊藿为狭披针形,基部位紫红色,其余部分为白色;巫山淫羊藿为披针形,颜色由浅黄变为黄色。粗毛淫羊藿花距为紫色、弯曲,距体与花柄垂直;巫山淫羊藿黄色,距多变,有直距、弯矩(向上、向下);宝兴淫羊藿为黄色,弯曲。宝兴淫羊藿花药黄色,粗毛淫羊藿花药为绿色,巫山淫羊藿黄药浅黄或浅绿色,三者花药均纵裂。胚珠数上粗毛淫羊藿为10~13粒;巫山淫羊藿为10~12粒;宝兴淫羊藿为9~12粒。图1A~C、图1D~F和图1G~I分别为宝兴淫羊藿、粗毛淫羊藿和巫山淫羊藿的单花及其开花过程。单花开花过程如下:第一天,内萼片对称的两片先展开,另两片随后开放,因相隔时间较短,4萼片基本同步展开并将外萼片撑开使其脱落;第二天,花朵距尖端分开但整体仍呈圆形,花药露出但仍被紧紧包裹;第三天,距尖离开并向外扩展,开始分泌花蜜,柱头开始伸长,花药开始散粉;第四天,萼片开到最大,距的伸展比内萼片更大,花药持续散粉;第五天,花朵完全展开,花蜜分泌量达到最大,花药颜色加深,花粉成熟粉散出,3~4 d后基本完全散出,雄蕊逐渐枯萎变成黑色并开始脱落,距萎奄,完成单花开花过程。
2.3 种群开花期
3种淫羊藿属植物均每年开花一次,三者始花日期、盛花期、终花期及花期持续时间具有差异。巫山淫羊藿的始花期最早,一般在3月下旬,种群开花历时22~27 d,单株花期为12~17 d,单花花期为4~5 d。粗毛淫羊藿始花期为3月下旬,种群的花期在22~33 d,单株开花时间一般为10 d左右,单花花期为4~5 d。宝兴淫羊藿始花期最晚在4月上旬,终花期也最晚,种群花期在25~40 d,单花花期为5~7 d。
2.4 访花者观察及实验
2.4.1 有效访花行为观察
通过访花观察及镜检发现3种淫羊藿属植物的访花者主要是膜翅目和双翅目昆虫(图2:A~F)。熊蜂为巫山淫羊藿的有效传粉者,以觅食花粉和花蜜为报酬,根据花距和喙的长短,不同种类熊蜂访花行为不同,熊蜂属1从距的中部打洞吸取花蜜其腹部为主要的传粉部位(图2:A);熊蜂属2从距开口处吸蜜,熊蜂颈部为主要的传粉部位(图2:B)。粗毛淫羊藿观察到的访花者有3种,只有熊蜂是有效传粉者,熊蜂从距的开口处吸食花蜜,期间花粉接触其腹部,在吸食花蜜的过程中完成传粉过程(图2:C)。宝兴淫羊藿观察到访花者有3种(图2:D~F),有效传粉者为蜜蜂科蜜蜂属中的隧蜂(图2:D),隧蜂从距的开口处钻进花内收集花粉,用中足自上而下收集花粉,期间刮落的花粉散落在其腹部。双翅目眼蝇科的拟蜂眼蝇(图2:E)和蚜蝇科食蚜蝇(图2:F),以取食花粉为目的,访花过程前足抓住雄蕊,后主抓住距的开口处,其口器伸出在雄蕊上取食花粉,期间头部、口器沾上花粉,以此同时头部及口器接触到宝兴淫羊藿柱头完成了传粉。
2.4.2 访花频率与花部特征诱物试验
3种大花类淫羊藿属植物的访花者主要集中在开花后第2~3 d,第4 d后访花者明显减少。阴雨天气访花者几乎没有,阴天有少量访花者,晴天访花者比较多,晴天时有效传粉者的访花频率对比情况如下(图3:A):7:30后开始有访花昆虫,访花频率最高出现在10:00~12:00和14:00~17:00时段,此时段光照强度和温度升高,湿度减小,有利于昆虫活动。3种植物材料的访花频率存在明显差异,其中,巫山淫羊藿的访花者数量最多。3种植物材料在开花第三天,花蜜量差异显著(P<0.05)(图3:B),访花频率的差异可能是因为巫山淫羊藿中距的花蜜酬物较多,而粗毛淫羊藿距中的花蜜酬物较少,宝兴淫羊藿传粉者主要以花粉为酬物。去除花距后,3种植物材料的花朵均有访花者去拜访,但数量显著低于自然状态下的访花数量,说明花距可能是淫羊藿属植物吸引访花昆虫的重要诱物,特别是巫山淫羊藿和粗毛淫羊藿,有效访花者主要以花距中的花蜜为酬物,而宝兴淫羊藿花距去除后访花者数量明显减少,可能是其黄色花距有吸引昆虫的作用,因此去除花距直接导致访花者数量降低。
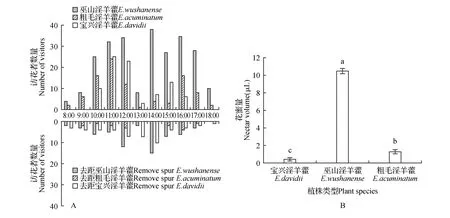
图3 有效访花者的访花频率及单花花蜜量Fig.3 Visiation frequency of effective pollinators and single flower nectar volume

表2 柱头可授性及结实率测定结果
注:1.自然对照组;2.套袋不处理;3.开花当天,同株自花授粉;4.开花当天,同株异花授粉;5.开花当天,去雄不套袋;6.开花当天,去雄异株异花授粉;7.开花第2天,去雄异株异花授粉;8.开花第3天,去雄异株异花授粉;9.开花第4天,去雄异株异花授粉;10.开花第5天,去雄异株异花授粉
Note:1.Natural control;2. Bagged control;3. Self-pollination;4. Same strain cross-pollination;5.Remove anthers and unbagged;6. First day of flowering dioeciously cross-pollination;7.Second day of flowering dioeciously cross-pollination;8.Third day of flowering dioeciously cross-pollination;9.Fourth day of flowering dioeciously cross-pollination;10.Fifth day of flowering dioeciously cross-pollination
2.5 柱头可授期及座果率
从表2可以看出,套袋及同株异花授粉均未见座果,说明3种淫羊藿均不能进行自花授粉;人工自花授粉结实率为0,说明其也不能进行无融合生殖;去雄不套袋处理中,巫山淫羊藿、粗毛淫羊藿和宝兴淫羊藿的结实率分别为58.33%、51.67%、35%,说明3种试验植物必需依靠媒介才能完成传粉过程。开花当天和开花第2 d人工授粉的结实率都较高,说明开花1~2 d时柱头可授性较强;开花第3~4 d授粉的结实率明显下降,表明此时柱头具有可授性,但有所降低;开花第5 d,3种试验材料的柱头均丧失可授性。另外,3种试验材料对照组的花朵,座果率大小情况为:巫山淫羊藿>宝兴淫羊藿>粗毛淫羊藿;与开花当天进行异株异花人工授粉的座果率相比明显降低,表明3种试验材料在开花期间有效访花传粉能力有限。在野外资源调查时,发现3种淫羊藿属植物存在“花多果少”的现象,估计与传粉者数量不足有关,特别是粗毛淫羊藿与宝兴淫羊藿花蜜量较少,吸引以访花昆虫能力不足。
2.6 生殖因子与环境因子相关性分析
3种淫羊藿属植物的生殖因子与环境因子具有一定相关性(表3),其中,巫山淫羊藿的开花期和温度与光照强度具有显著负相关,粗毛淫羊藿和宝兴淫羊藿的开花期与温度呈显著负相关,说明温度是影响淫羊藿属植物花期的显著因子,温度越高其开花持续时间就越短。花蕾数中巫山淫羊藿与温度和光照强度呈显著正相关,宝兴淫羊藿中与温度显著正相关,这说明在相对较高的温度下,3种淫羊藿属植物的个体开花数量较高。访花频率中3种淫羊藿属植物均与温度呈显著或极显著正相关,巫山淫羊藿与粗毛淫羊藿中的访花频率与光照强度呈显著相关,这说明温度和光照强度是影响访花者进行访花活动的重要影响因子。自然座果率中3种淫羊藿属植物均与温度呈显著正相关,巫山淫羊藿和粗毛淫羊藿均与光照强度显著正相关,说明3种淫羊藿属植物的生殖特征与生境中的温度和光照强度具有密切的联系。

表3 生殖因子与环境因子的相关性
注:**极显著水平为0.01;*显著 1.巫山淫羊藿;2.粗毛淫羊藿;3.宝兴淫羊藿
Note:**was lextremely significant;*was significant 1.E.wushanense; 2.E.acuminatum; 3.E.davidii
3 讨论
3.1 花部特征、繁殖系统对淫羊藿生殖成功的影响
繁殖系统控制种群繁殖的各种生理和形态机制,在决定植物的进化路线、基因流和表征变异上起着重要作用。花粉通过媒介(风力、昆虫、鸟等)移动到柱头中,这一过程的前提是柱头必须具有可授性[17]。人工授粉不同组合试验表明,3种淫羊藿自花授粉不育,为虫媒异交植物且自交不亲和,柱头可授期为开花第1~4 d,在第2 d进行授粉效果最佳。花性状主要包括花的颜色、形状、大小、对称性、花粉数量和颜色、花粉胚珠比等,其主要功能是为了吸引传粉者提高异交繁殖[18~19]。不同物种中由于花部特征差异而形成不同类群的传粉者,而传粉者又反过来促进不同物种的花部特征演化[20~23],本研究中3种淫羊藿虽同属于大花类群,但其花部特征完全不同,因此具有不同类群的传粉者。巫山淫羊藿的花距具有3种形态:直距、距尖向上和距尖向下,访花者中熊蜂属1趋向访问距尖向上的花朵,而熊蜂属2则趋向访问距尖向下和直距的花朵,这可能是巫山淫羊藿花部特征与访花者的协同进化的结果。巫山淫羊藿的自然座果率最高,各个时间段里有效访花者的数量也是最高的,而宝兴淫羊藿和粗毛淫羊藿传粉者数量远比巫山淫羊藿的数量少,因此,粗毛淫羊藿及宝兴淫羊藿的自然座果率略低,不同的淫羊藿属植物其结实特性存在显著差异,这与Suzuki等人研究结果一致[24]。不同淫羊藿的访花者的访花时间、访花长短和访花频率不一致,如没有传粉昆虫,将会严重影响淫羊藿属植物有性繁殖的成效。在移植和栽培淫羊藿属植物时,要充分考虑传粉者的生存条件,开花期间提供条件吸引有效传粉者。诱物试验表明3种淫羊藿植物传粉者最主要的诱物是产生花蜜的距,其他部位,如萼片颜色、距的长度和颜色等对生殖特征有无影响还待进一步研究。
3.2 生殖特征与生境因子关系
淫羊藿属植物的生殖活动一般在3~5月份,此时水分充足且温度适宜,同时大多数传粉者的生殖活动也在这个时候。植物间为竞争传粉者,往往会采取"集中,大量开花模式"获得传粉者的有效访问,达到生殖成功的目的。不同区域植物开花时间的差异与环境因子(如光照、温度和湿度)的变化具有显著相关性[25~26]。3种淫羊藿属植物均为集中大量开花模式,巫山淫羊藿的始花期最早,相关性分析表明,温度和光照是重要的生态限制因子,这与权秋梅的研究结论一致[6]。粗毛淫羊藿在峨眉山多雾多雨的3月下旬~5月上旬进入花期;宝兴淫羊藿所处海拔位置较高,温度较低,始花期最晚,同时花期也较长,有利于提高传粉机会。3种淫羊藿属植物的自然座果率均与温度呈显著正相关,巫山淫羊藿和粗毛淫羊藿均与光照强度呈显著正相关,说明3种植物的生殖特征与温度和光照强度相关。因此,对淫羊藿属植物进行有性繁殖或栽培时,应充分考虑到生境的异质性,提供适宜的光照和温度条件。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
支部建设(2019年24期)2019-09-28
重庆与世界(2019年9期)2019-09-10
金桥(2019年12期)2019-08-13
文苑(2019年14期)2019-08-09
火花(2018年5期)2018-11-21
中国报道(2018年4期)2018-06-11
大科技·百科新说(2018年3期)2018-04-11
山东青年(2016年3期)2016-02-28
