
秦王川盐沼湿地芦苇叶片比叶面积与光合效率的关联分析
2019-11-13 00:39:14赵成章赵连春王继伟
生态学报 2019年19期
李 群, 赵成章, 赵连春, 王继伟, 文 军
西北师范大学地理与环境科学学院, 甘肃省湿地资源保护与产业发展工程研究中心, 兰州 730070
植物是一个功能和结构平衡的生命体,在长期的进化和自然选择过程中,植物通过调整内在和外在结构优化其功能和结构以适应周围环境变迁[1- 2]。植物叶片的结构性状在一定程度上决定叶片光合生理活性,能够反映植物叶片对所吸收光能的利用水平[3],叶绿素荧光与叶片的光合作用效率密切相关[4],可反映植物的光合效率及在逆境条件下的生理状态。光合作用即植物叶片将吸收的光能转化为化学能的过程,包括光系统(PSI和PSII)[5],当植物受到环境胁迫时,植物往往形成较低比叶面积(SLA, specific leaf area)的叶片,通常用于构建保卫结构、叶肉细胞密度和增加叶片厚度的碳同化产物比例居多,有助于防止叶片散失水分和提高叶片的光合效率[6];另一方面,叶绿素荧光中的非光化学淬灭在保护植物光系统II免受光抑制及光损伤方面发挥重要作用,能够通过降低电子传递速率,致使光化学活性下降,从而引起光合速率下降[7],由此表明比叶面积与叶片光合效率关系十分密切。盐胁迫被广泛的描述为影响植物生长和发育的重要环境因子之一[8]。盐胁迫使植物吸水受阻,碳同化速率下降,在土壤水盐差异条件下,为适应环境,植物需要对叶片碳同化产物在叶厚度、比叶面积等叶片性状上权衡分配,并通过调整叶片电子传递速率和光能分配等途径改变叶片光合作用,提高植物对异质性环境的适应能力,实现植物对叶片吸收光能的有效利用,这体现了植物叶性状与叶片光合和荧光生理特性间的协同适应机制。因此,通过分析秦王川国家湿地公园典型盐沼湿地湿生植物叶片比叶面积-实际光合效率,比叶面积-电子传递速率的相互关系,有助于探究芦苇叶片比叶面积与光合效率的内在关联性和对盐沼湿地特殊生境条件下植物光合产物的积累与分配间权衡策略的认识和理解,揭示了盐沼湿地植物适应特殊生境所形成的生态适应机制的科学意义和理论价值。
芦苇(Phragmitesaustralis)属于多年生根茎型禾本科植物,具有极强的环境适应能力,广泛分布于西北干旱内陆盐沼湿地和沼泽湖泊区,也是西北干旱区盐沼湿地生态系统的优势植物[9]。芦苇具有适应性强、生长范围广、经济价值高、生态功能多样等多种优势[10],而国内外有关芦苇的研究主要集中在芦苇叶片光合作用对土壤水盐因子的响应[11]、叶片比叶面积与热耗散关联性分析[12]、对胁迫生境的适应机制[13]等方面;很少从芦苇叶性状与光合、荧光特性出发分析其特殊的环境适应机制,鉴于此,我们在实地考察的基础上,以植物叶性状相关理论为基石,研究了秦王川国家湿地公园典型盐沼湿地湿生植物芦苇叶片比叶面积-实际光合效率,比叶面积-电子传递速率的相互关系及变化规律,试图明晰:(1)盐沼湿地芦苇SLA与叶片光合效率间存在何种关联性?(2)形成这种关系的主要原因有哪些?旨在丰富对盐沼湿地植物长期适应环境形成的生存策略的认识和理解。
1 研究地区与研究方法
1.1 研究区域和样地概况
研究区位于甘肃省兰州市的秦王川盆地(图1), 地理坐标为36°25′ N—36°29′ N,103°37′E—103°41′E, 海拔1895—1897 m, 属大陆性季风气候, 年平均气温6.9 ℃, 年降水量220 mm, 主要集中在6—9月, 年日照时间2700 h, 平均无霜期126d。研究区地处乌鞘岭褶皱山岭南侧的边缘低山区,四周山地环绕,中心地带地势平坦开阔,形成盆地,盆地南部广泛分布第四系松散层孔隙潜水,含水层为砂碎石及中细砂层,受常年或季节性水淹的影响,土壤大部分以非盐渍土和轻盐渍土为主,发育了水生和湿生植物群落,以湿生和挺水植物为主。主要植物有:芦苇(Phragmitesaustralis)、枸杞(Lyciumchincnse)、多枝柽柳(Tamarixramosissma)、香蒲(Typhaangustifolia)、灰绿藜(Chenopodiumglaucum)、紫菀(Astertataricus)、假苇拂子茅(Calamagrotispseudophragmites)、碱蓬(Suaedasalsa)、苦苣菜(Sonchusoleraceus)、盐角草(Salicorniaeuropaea)、早熟禾(Poaannua)等。
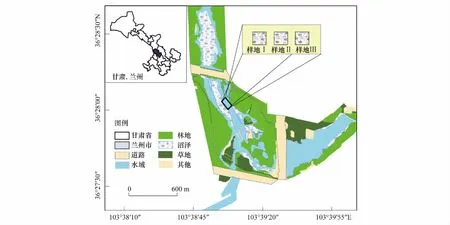
图1 研究区概况和样地设置图Fig.1 Study area and locations of measured plots
1.2 实验方法
于2016年9月,依据前期对土壤盐分分布规律和土壤含水量的调查,我们发现试验地土壤含水量无显著差异(表1),土壤盐分差异显著(表2, 表3),据此在距离水域远近形成的天然盐分梯度的基础上选择一块由芦苇群落末端向水域边缘平缓延伸,面积约100 m×100 m,芦苇为单一优势种的草本沼泽盐沼湿地进行实验。首先按盐分梯度的划分,从芦苇群落末端至水域边缘,间隔5 m依次布置3条宽20 m×20 m的样带,进行土壤盐分的测定(在每条样带上从远水区开始每间隔5 m采集表层土壤,测定土壤电导率)。根据土壤盐分变化规律,在3条平行样带上从芦苇群落末端向水域边缘依次设置3个10 m×10 m的样地Ⅰ、样地Ⅱ和样地Ⅲ,分别为:(1)样地Ⅰ位于远水区芦苇群落末端,土壤表层有盐碱累积,芦苇生长在该生境中,植被低矮密度较大,土壤电导率介于2.3—2.8 ms/cm;(2)样地Ⅱ位于中水区属盐渍化和非盐渍化土壤的过渡地带,芦苇生长在该生境中,植被高度和密度属于过渡型,土壤电导率介于1.8—2.2 ms/cm;(3)样地Ⅲ位于水域边缘,季节性淹水,土壤不存在盐渍化,该生境下芦苇植株高大,密度小,土壤电导率介于0.8—1.5 ms/cm;接着按已划分的芦苇种群的盐分梯度样地,每个样地依次设置6个2 m × 2 m样方,总计18个样方。
首先分别测定每个样方内湿地群落所有植物的高度和密度,芦苇以株为单位;其次在每一个样方随机选择大中小各两株,总计6株芦苇,先用卷尺测量株高(从基部到最高分枝的距离);然后选取叶片,从每株芦苇的顶端、中间、底层的最外层选取充分伸展且健康完整的3个叶片,用于芦苇叶性状的调查:
(1)光合特征值测定:选择晴朗天气的9:00—12:00对标记过的芦苇进行植物气体交换等参数的测定,光合测定使用GFS- 3000便携式光合测量系统(Heinz Walz GmbH, Bavaria, Germany),测量过程中使用人工红蓝光源,光合有效辐射(PAR)为1200 μmol m-2s-1,CO2浓度约为340 μmol;/mol,流速设定为750 μmol/s,相对湿度(RH)保持在60%—70%,叶温(Tleaf)保持在15—20 ℃。选取每个植株中端展开的4片成熟健康叶片分别测定叶片的净光合速率(Pn)、蒸腾速率(Tr)等参数,待数据稳定后读取数据。
(2)芦苇种群PAR的测定:选择与光合参数测定同一天,用手持光量子计(3415F, 3415F, Spectrum technologies, Aurora, USA)于9:00—11:00对3样地内芦苇种群的上方(植株顶端10 cm)、中层(介于上方和地表的中间)和地表(距离地面10 cm)处进行PAR测定,重复3次,计算平均值[12]。
(3)叶面积(LA)、叶干重(LDW)和比叶面积(SLA)测定:将测定完光合作用的叶片装入带有编号的信封迅速带回实验室,用便携式激光叶面积仪(CI- 202, Walz, Camas,USA)扫描每个叶片,相应的叶面积数据导出用于计算比叶面积;最后将叶片再次装入信封,于80 ℃烘箱中烘干48 h,称叶干质量即叶干重(精度0.0001g)。SLA=LA/叶干质量(LDW)。
(4)叶厚度测定:使用游标卡尺测量(精确到0.01 mm),测量时避开叶脉,并尽量选择叶片中心部位,每个个体在不同部位测量5次,最后取平均值为该个体的叶片厚度。
(5)土壤含水量测定:每个小样方随机挖掘1 m×1 m×0.5 m土壤剖面,用环刀(200 cm3)分5层间隔10 cm采取土样,重复3次,各土层均匀混合,现场编号装入铝盒称鲜量后带回实验室在105 ℃的烘箱内烘12 h,取出称重,计算出各样地土壤含水量。
(6)土壤盐分测定:用电导法测定土壤含盐量,在室温下称取过2 mm筛的风干土样10 g,加50 mL去CO2蒸馏水(水土质量比为5∶1),取浸出液,置振荡机上振荡5 min。将布氏漏斗与抽气系统相连后把悬浊的土浆缓缓倒入漏斗,直至抽滤完毕,滤液倒入三角瓶备用[14]。用上海雷磁仪器厂生产的DDS- 11C便携式电导仪测浸出液的电导率,3次重复,取平均值。
(7)叶绿素荧光参数测定:对各样地芦苇叶片现场进行叶绿素荧光参数测定,采用调制式IMAGING-PAM叶绿素荧光仪(Heinz Walz GmbH, Effeltrich, Germany)测定叶绿素荧光参数,手动设置光化光强度为1200 μmol m-2s-1,叶片先经暗适应30 min,测定初始荧光产量(F0)和最大荧光产量(Fm),随后打开光化光,在软件的Kinetics窗口检测各叶绿素荧光参数的荧光诱导曲线,相应的数据可直接从Report窗口导出[12]。相关的叶绿素荧光参数为最大光合效率(Fv/Fm),光化学淬灭系数(QP),非光化学猝灭系数(NPQ),非调节性能量耗散的量子产量(Y(NO)),调节性能量耗散(Y(NPQ)),实际光合效率(Y(II)),电子传递速率(ETR),具体操作参考Schreiber等[15]和Schreiber[16]的方法;
①Fv/Fm=(Fm-F0) /Fm. (Fv, 饱和脉冲光导致的荧光产额)
②QP=(Fm′-Fs) /Fv′=1-(Fs-Fo′) / (Fm′ -Fo′). (Fm′, 光下最大荧光;Fo′, 光下最小荧光;Fv′, 饱和脉冲光导致的荧光产额;Fs, 稳态荧光)
③ NPQ=(Fm-Fm′)/Fm′=Fm/Fm′-1.
④ PSII实际光合效率Y(II) = ΦPSII=(Fm′-Ft)/Fm′. (Ft, 实时荧光)
⑤ PSII非调节性能量耗散的量子产量Y(NO)=1/(NPQ+1+qL(Fm/Fo-1)). (qL, 基于“湖泊模型”的光化学淬灭系数)
⑥ PSII调节性能量耗散的量子产量Y(NPQ)=1-Y(II)-Y(NO).
⑦ ETR = 0.5 ×Y(II) × PAR × 0.84 (μmol m-2s-1). (PAR, 光合有效辐射)
1.3 数据处理
实验所得所有数据运用SPSS 20.0 (SPSS Inc., Chicago, IL, USA)进行相关分析和单因素的方差分析(ANOVA)。各功能性状间做回归分析,相关性最高用做定量分析。数据分析主要采用SMA的方法[17],首先对所有样地芦苇的比叶面积、ETR和Y(II)进行统计分析,对分析数据进行以10为底的对数转换,然后采用y=axb,线性转换成log(y)=log(a)+blog(x),式中,x和y表示两个特征参数,b代表性状关系的截距,a表示相关性的斜率,即异速生长参数或相对生长的指数,当a=1时,表示两者是等速关系;当a>1时,表示y的增加程度大于x的增加程度;当a<1时,表示y的增加程度小于x的增加程度。
2 结果与分析
2.1 湿地群落生物学和土壤含水量特征
各样地群落生物学特征与土壤含水量呈现规律性梯度变化,表现出不同程度的差异(表1)。不同样地群落的密度、高度、PAR、地上生物量差异性均显著(P<0.05),从样地Ⅰ到样地Ⅲ,群落高度和地上生物量呈增加趋势,分别增加了1.32和1.94倍;而群落密度呈先增加后减小的倒U型趋势,减少了51.15%;PAR呈减少趋势,减少了52.52%;土壤含水量差异不显著,呈增加的趋势,增加了33.40%,由于研究地浅层地下水及地势等因素的影响,土壤含水量充足,均大于30%,植被生长不存在水分短缺的限制。
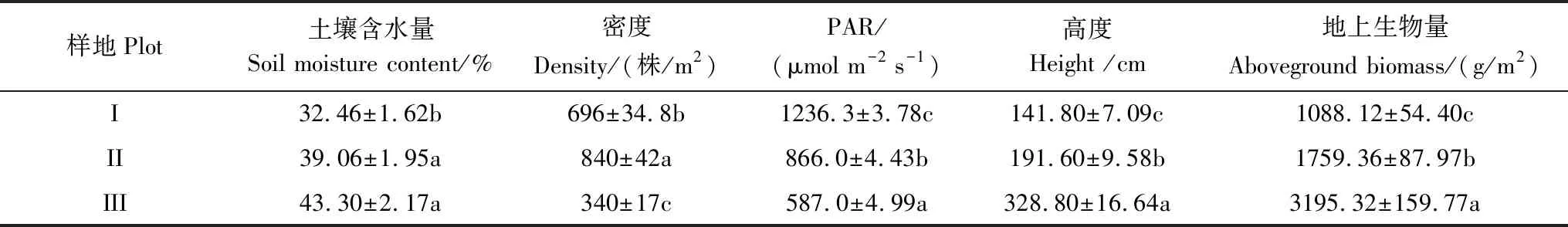
表1 不同样地的湿地群落生物学特征和土壤含水量(数值为平均值±标准误差)
PAR: 光合有效辐射, Photosynthetic active radiation;同列不同小写字母表示样地间差异显著(P< 0.05)
2.2 研究区土壤盐分特征
参照相关土壤盐渍化分级标准[18],采样区域内土壤含盐量分为3级:4—8 ms/cm,属重盐渍土;2—4 ms/cm,属中盐渍土;1.8—2 ms/cm,属弱盐渍土。
2.2.1研究区土壤盐分含量分析
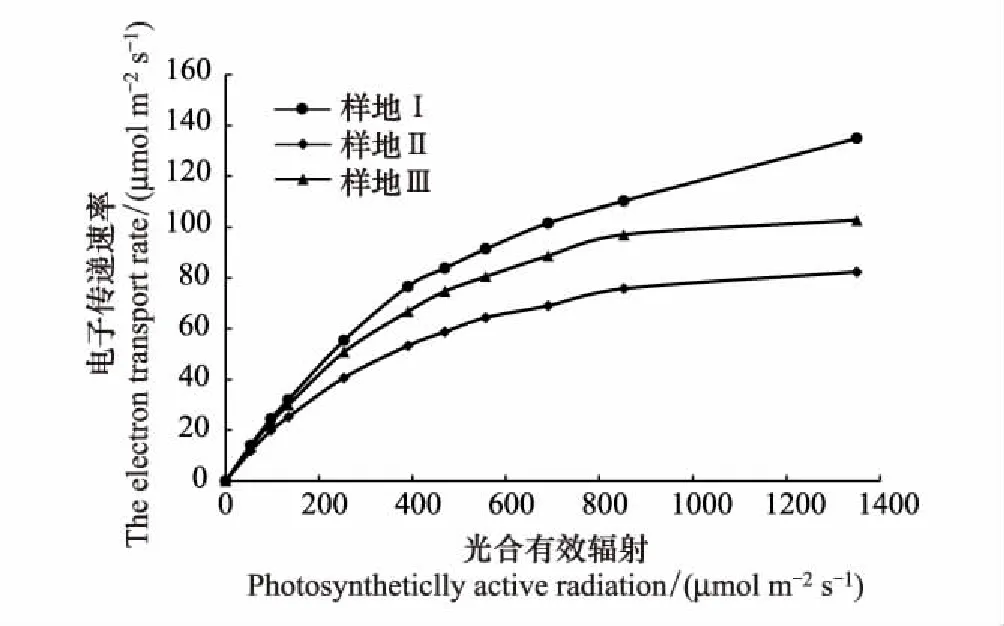
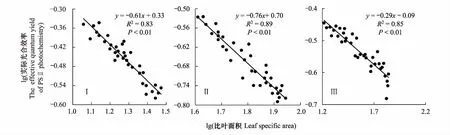
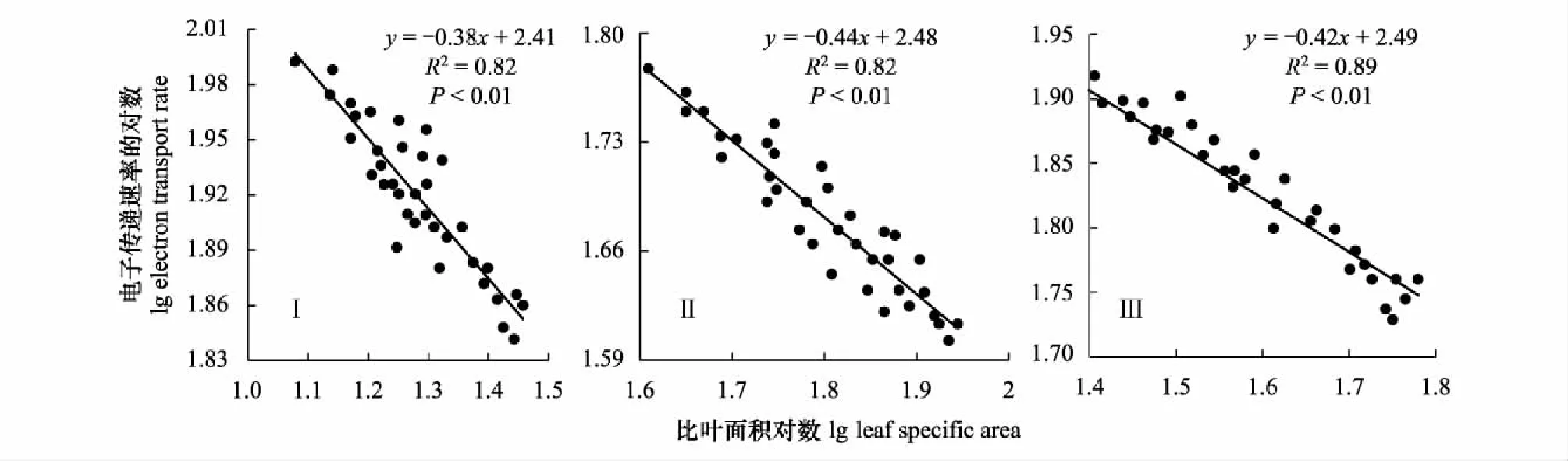
由表2可以看出,土壤含盐量的平均值为2.01 ms/cm,中值为2.04 ms/cm,二者差异不大,中值略大于平均值,说明土壤含盐量基本呈正态分布,因此采用平均值作为衡量研究区土壤盐渍化指标较为适宜,研究区土壤属中度盐渍化水平。变异系数(C.V)是衡量测定值在空间上离散和变异程度的指标,土壤含盐量变异系数为0.31,表现为在水平方向上呈中等变异性(根据变异系数的划分等级:C.V>1为强变异性或数据离散程度高,0.1 表2 研究区土壤盐分统计特征 2.2.2各样地土壤盐分含量及垂直分布特征 由表3可以看出,3个样地土壤盐分含量(0—30 cm土层)均值波动范围为1.30—2.58 ms/cm,样地Ⅰ土壤盐分含量均值为2.58 ms/cm,样地Ⅲ土壤盐分含量均值为1.30 ms/cm,极差达1.28 ms/cm,其变化趋势为:样地Ⅰ>样地Ⅱ>样地Ⅲ,样地间差异显著(P<0.05),与盐渍化生境有关。各样地20—30 cm土层的盐分含量呈现不同的差异性,样地Ⅲ和样地Ⅱ不同土层间的盐分含量均存在显著差异(P<0.05);样地Ⅰ不同土层间的盐分含量差异均不显著,20—30 cm土层与其他土层间差异显著(P<0.05)。 表3 各样地土壤盐分含量特征 同行不同小写字母表示样地内差异显著(P< 0.05),同列不同大写字母表示样地间差异显著(P< 0.05) 根据实验测量调查分析,芦苇叶性状和光合参数特征见表4。单因素方差分析表明,不同土壤盐分条件下,芦苇叶性状间、光合参数特征间呈显著差异(P<0.05)。随着土壤盐分含量的降低,芦苇叶面积、株高、Pn、Tr和WUE呈逐渐增大的趋势,分别增大了2.73、1.32、1.17、0.17倍和0.86倍;从样地Ⅰ到样地Ⅲ,叶厚度减小了25%;叶干重呈先减少后增大的U型变化趋势,整体增大了0.85倍;比叶面积呈先增大后减小的趋势,整体增大了1.02倍。表明与土壤盐分含量高的样地Ⅰ相比土壤盐分含量最小的样地Ⅲ更适合芦苇植株的生长。 表4 不同样地芦苇叶性状与光合生理参数特征(数值为平均值±标准误差) Pn,: 净光合速率, net photosynthetic rate;Tr: 蒸腾速率, transpiration rate; WUE: 水分利用效率, water use efficiency;同行不同小写字母表示样地间差异显著(P< 0.05) 根据野外实验测量和分析,芦苇叶片荧光参数特征见表5。单因素方差分析表明:不同土壤盐分条件下,芦苇叶绿素荧光特征值间呈显著差异(P<0.05)。随着土壤盐分含量的降低,芦苇的Y(II)和 ETR呈先减小后增大的趋势,整体减小了17%和16%;QP呈先减小后增大的趋势,整体无显著变化;NPQ和Y(NPQ)呈先增大后减小的趋势,整体减小了37%和18%;Y(NO)增大了0.45倍;Fv/Fm呈先增大后减小的趋势,整体减小了32%。表明与样地Ⅰ和样地Ⅱ相比样地Ⅲ中芦苇叶片用于热耗散的能量在降低,而以荧光的形式消耗的能量在增加,体现了芦苇植株为适应特殊生境,适时调整PSII天线色素吸收光能的分配以实光合效率最大化,维持自身的生长和繁殖。 不同土壤盐分条件下芦苇叶片中PSII电子传递速率快速光响应曲线存在显著差异(P<0.05, 图2),单因素方差分析表明,随着土壤盐分含量的降低和光照强度的增加,芦苇叶片的ETR呈样地Ⅰ > III > II的变化趋势,且当饱和脉冲光强大于1050 μmol m-2s-1时,各样地芦苇叶片的ETR逐渐由增加趋势转为平稳的趋势。表明在光照强度一定的条件下,土壤盐分含量直接影响了芦苇叶片的ETR,进而影响芦苇植株的光合作用,且由ETR样地Ⅰ >III > II,可说明土壤盐分在一定程度上可促进叶片的电子传递。 表5 不同样地芦苇叶片荧光参数特征(数值为平均值±标准误差) Y(II): 实际光合效率, The effective quantum yield of PSII photochemistry;QP: 光化学淬灭系数, Photochemical quenching; NPQ: 非光化学猝灭系数, Non-photochemical quenching;Y(NO): 非调节性能量耗散的量子产量, Quantum yield of non-regulatory energy dissipation;Y(NPQ):调节性能量耗散的量子产额, The quantum yield of regulatory energy dissipation; ETR: 电子传递速率, Electron transport rate;Fv/Fm, 最大光化学效率, The maximum efficiency of photosystem (PS) II photochemistry;同列不同小写字母表示样地间差异显著(P< 0.05) 图2 不同样地芦苇叶片暗适应后电子传递速率的快速光响应曲线Fig.2 Light-response curve of ETR (apparent electron transport rate) in the dark-adapted leaf samples of reed under different plots 快速光响应曲线(RLC)是在一系列间隔20s的光强(0, 150, 300, 450, 600, 750, 900, 1050, 1350 μmol m-2 s-1)下获得的;数据重复3次平均值 如图3所示,根据标准化主轴估计分析,芦苇SLA与Y(II)的相关性在样地Ⅰ、样地Ⅱ和样地Ⅲ均呈极显著负相关关系(yI=-0.61x+0.33,R2=0.83,P<0.01;yII=-0.76x+0.70,R2=0.89,P<0.01;yIII=-0.29x-0.09,R2=0.85,P<0.01);芦苇的SLA与Y(II)的SMA斜率的绝对值呈先增大后减少的变化趋势,其中在土壤盐分含量高的样地Ⅰ为-0.61 (95%的置信区间CI=(-0.34, -0.35)),样地Ⅱ为-0.76 (95%的置信区间CI=(0.24, 0.25)),样地Ⅲ为-0.29 (95%的置信区间CI=(0.28, 0.29)),且均与-1存在显著差异(P<0.05)。表明随着SLA的增加,Y(II)呈减少的趋势,不同土壤盐分含量条件下芦苇的SLA和Y(II)呈“此消彼长”的权衡关系,其中土壤盐分含量居中的样地Ⅱ,SLA最大,Y(II)最小。表明在土壤盐分含量差异条件下,芦苇会选择不同的SLA和Y(II)的特征组合,以实现碳同化产物的最优分配和对叶片吸收光能的有效利用。 图3 不同样地芦苇实际光合效率(Y(II))与比叶面积(SLA)之间关系Fig.3 Relationship between the effective quantum yield of PSII photochemistry (Y(II)) and specific leaf area (SLA) of Phragmites australis among different plot 如图4所示,根据标准化主轴估计分析,芦苇SLA与ETR的相关性在样地Ⅰ、样地Ⅱ和样地Ⅲ均呈极显著负相关关系(yI=-0.38x+2.41,R2=0.82,P<0.01;yII=-0.44x+2.48,R2=0.82,P<0.01;yIII=-0.42x+2.49,R2=0.89,P<0.01);芦苇的SLA与ETR的SMA斜率的绝对值呈先增大后减少的变化趋势,其中在土壤盐分含量高的样地Ⅰ为-0.38 (95%的置信区间CI=(0.33, 0.34)),样地Ⅱ为-0.44 (95%的置信区间CI=(0.24, 0.25)),样地Ⅲ为-0.42 (95%的置信区间CI=(0.27, 0.28)),且均与-1存在显著差异(P<0.05)。表明随着叶片SLA的增加,ETR呈减少的趋势,3个样地的芦苇SLA和ETR均呈“此消彼长”的权衡关系,土壤盐分含量居中的样地Ⅱ,SLA最大,ETR最小。表明面对不同土壤盐分含量的生境,芦苇会选择不同程度的叶片ETR和改变叶片SLA的方式,实现光合产物的合理分配和高效的光合效率来维持自身的生长和繁殖。 图4 不同样地芦苇电子传递速率(ETR)与比叶面积(SLA)之间关系Fig.4 Relationship between electron transport rate (ETR) and specific leaf area (SLA) of Phragmites australis among different plot 光合作用是绿色植物利用光能,把CO2和H2O合成有机物,同时释放O2的生理过程[19],叶绿素荧光与光合作用密切相关,通过分析叶绿素荧光可探究光合作用的过程[20]。研究发现,在3种土壤盐分含量的生境条件下,SLA与实际光合效率(Y(II))和ETR均呈极显著负相关关系(P<0.01),反映了土壤盐分对湿地植物叶片构型和光合效率的影响及湿地植物对特殊生境的适应机制。 SLA作为叶片结构的重要特征之一,也是影响WUE的重要因素[21],能够反映植物在异质环境条件下的水分的利用效率和自我保护能力[22]。湿地群落土壤盐分含量最大的样地Ⅰ,群落高度和地上生物量最小,密度较低(表1),PAR和土壤盐分含量最大(表1, 表3),叶片ETR表现为最大(表5, 图2),SLA较低(表4),SLA与Y(II)和ETR间形成了极显著的负相关关系(P<0.01, 图3, 图4)。这主要是因为:(1)该生境芦苇植株间邻体干扰和遮光现象比较弱,不仅光照强烈,土壤盐分含量也最大(表3),使得芦苇面临水盐双重胁迫的环境压力,在资源有限的情况下,芦苇植株选择小叶模式,形成低SLA的叶片不仅可以增加叶片保水性,而且可以减缓叶温升高的速率,有利于延长叶片碳积累的时间和提高植物对有限水分的利用效率[23],因而在样地Ⅰ,芦苇的净光合速率、蒸腾速率和WUE表现为最低(表4);另一方面,强光条件下,芦苇叶片ETR的增加(表5, 图2)有利于其实现自我保护,这是因为植物光合机构通过电子流进行能量耗散是进行光保护的有效途径[24]。因而,SLA与ETR间形成了极显著的负相关关系(P<0.01, 图4)。(2)盐胁迫和强光条件下形成小而厚的叶片具有较高的热交换能力[25],在叶片吸收光能一定时,芦苇将更多的光能分配到光化学反应中心,因而Y(II)的值最高(表5),调节性能量耗散Y(NPQ)和非光化学淬灭系数(NPQ)较大(表5),必然要降低非调节性能量耗散(Y(NO)),合理分配叶片吸收的光能,从而实现叶片自我保护和光合作用的正常进行,这与武传兰等[7]的研究“盐胁迫下,植物叶片增加光化学反应中心对吸收光能的分配比例和非光化学反应耗散的能量,以维持植物正常的生理需求”结论相一致。因此,在土壤盐分含量高的样地Ⅰ,SLA与Y(II)间形成了极显著的负相关关系(P< 0.01, 图3)。体现了在胁迫生境下,植物通过调整叶片构型实现叶片吸收光能的合理分配以维持自身生长和繁殖的生存策略。 植物在异质生境中会通过调整植物形态结构的表型可塑性及资源的权衡分配来增加对环境适应能力[26- 27]。在土壤盐分含量最低的样地Ⅲ,湿地群落高度和地上生物量最大,密度最小(表1),植株之间存在遮阴现象,PAR最低(表1),Y(II)和电子传递速率较样地Ⅰ有所降低(表5, 图2),土壤含水量充足,含盐量最低(表1, 表3),芦苇株高最大(表4),SLA较高(表4),Y(NPQ)最小,Y(NO)最大(表5),SLA与Y(II)和ETR之间形成了极显著的负相关关系(P<0.01, 图3, 图4)。主要原因如下:(1)该样地中芦苇的光照强度最低,株高和地上生物量最大(表1),邻体间的遮阴现象使得植株间产生激烈的光竞争,为使碳同化速率最大化,芦苇增加叶片面积,降低叶厚度形成大而薄的叶片,获得较高的SLA(表4),捕光面积的增加使得净光合速率随之增大,从而实现了高水分利用效率(表4);另一方面,该样地芦苇种群植株底部的光照最弱(表1),因而叶片光合活性和叶片ETR不及样地Ⅰ (表5, 图2),SLA与ETR之间形成了极显著的负相关关系(P<0.01, 图4)。(2)样地Ⅲ大而薄的叶片厚的叶边缘阻止了叶片与周围的空气间的热量交换[28],使得用于叶片吸收的光能用于热耗散部分降低,光化学反应的部分增加,Y(II)和光化学淬灭(QP)表现为较大,NPQ和Y(NPQ)最小(表5);另一方面,样地Ⅲ中土壤含盐量最小(表3),芦苇不存在盐胁迫,生存环境良好,光照弱使得叶片的过剩光能少,因而芦苇叶片将更多的光能分配给了光化学反应和非光化学反应,减少了热耗散的能量分配,所以Y(NPQ)最低,Y(NO)随之增加(表5),(Y(II))较样地Ⅰ有所降低(表5),这与“叶片吸收的光能主要用于光合作用和叶片热耗散过程,因此二者是此消彼长的关系[29]”的结论一致。因而,SLA与Y(II)之间形成了极显著的负相关关系(P<0.01, 图3)。表明当植物叶片吸收光能一定时,必然会产生能量在光化学反应和非光化学反应之间的权衡分配,由此产生最有利于植物自身生长和存活的环境适应能力。 植物的叶片表皮会首先对环境变化做出响应[30],植物叶片荧光动力学测定的参数,可用于分析环境因子影响下植物叶片光合机构的变化[31]。样地Ⅱ土壤含盐量的降低,湿地群落高度、PAR、地上生物量均处于过渡区,密度最大(表1),若单纯选择小叶或者大叶模式,则不利于芦苇植株在有限资源下光合碳获取能力的最大化[32],所以芦苇选择了适中的叶面积和叶厚度,降低了叶片干物质积累使光合效率最大化,因而该样地叶片SLA最大,最大光化学效率(Fv/Fm)最大(表4, 表5),光照强度相比于样地Ⅰ有所降低,这是因为土壤盐分的降低使得芦苇植株吸水阻力减少,水分条件较样地Ⅰ有所改善,种群密度增大的结果,因此该样地叶片ETR表现为最低(表5, 图2);此外,与样地Ⅰ相比土壤盐分含量有所降低(表3),芦苇植株面临的盐胁迫得到了一定程度的缓解,生存环境得到改善,相比样地Ⅰ芦苇一方面增加干物质积累,Pn、Tr和WUE也有所增加(表4),另一方面,在叶片吸收光能一定的条件下,光化学反应消耗的能量表现为最低,而非光化学的能量有所增加(表5),这可能是因为虽然较样地Ⅰ土壤盐分含量、光照强度有所降低,但是芦苇植株光合活性和效率不及样地Ⅲ所致,所以Y(II)反而比样地Ⅰ低,导致SLA与Y(II)和ETR间仍呈现极显著的负相关关系(P<0.01, 图3, 图4),这体现了盐沼湿地植物随环境条件的改变,适时调整自身叶片构型及叶片吸收光能的分配,以寻求最佳的生存策略。 植物叶性状构建模式与光合效率的关系分析能反映植物在长期胁迫生境下形成的光合作用机理和特殊生存机制。生长在高盐分含量生境的芦苇,因为强光、高盐和缺水等胁迫生境迫使其选择小叶-低比叶面积的叶片构建模式,有利于实现异质生境下碳同化产物的最优分配和叶片对过剩光能的耗散;生长在土壤盐分含量低的生境下的芦苇植株,随着强光、水盐胁迫等的消失伴随的生存环境的改善,进而出现由于植株高大遮光严重引起的植株间光竞争现象,迫使其选择大叶-高比叶面积的叶片构建模式,不仅能够改善遮阴环境下植物吸收光能不足的问题,而且实现了光合收益的最大化和植物叶片对有限光能的合理分配。叶片比叶面积与光合效率的关系反映了特殊生境条件下湿地植物的叶片可塑性调节机制和特殊生存策略。
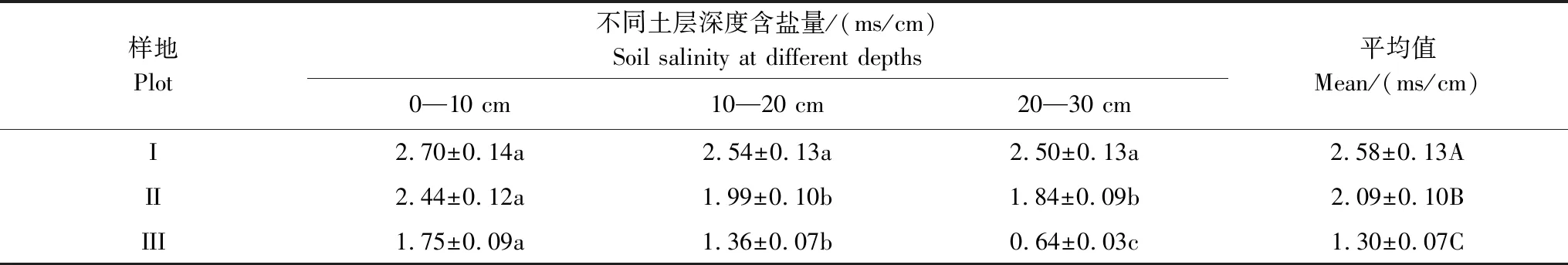
2.3 芦苇叶性状和光合参数特征
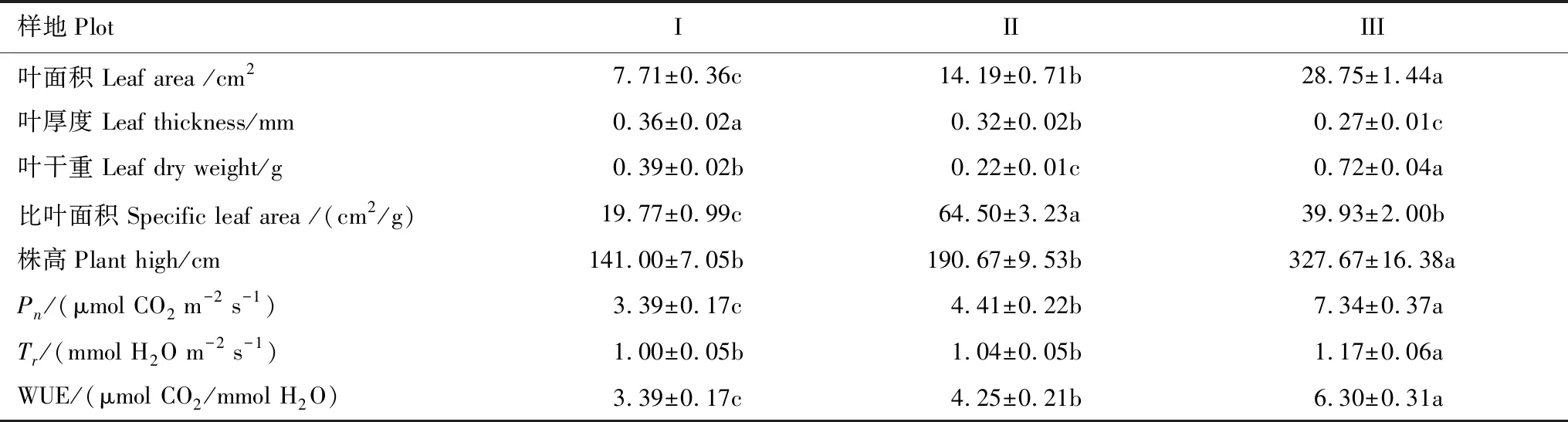
2.4 芦苇叶片荧光参数特征
2.5 不同样地芦苇叶片电子传递速率(ETR)的变化

2.6 芦苇SLA与实际光合效率(Y(II))的关系
2.7 不同样地芦苇SLA与电子传递速率(ETR)的关系
3 讨论
4 结论
猜你喜欢
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
落叶果树(2021年6期)2021-02-12 01:29:10
新疆农业科学(2020年1期)2020-02-14 03:22:46
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
读者·校园版(2016年6期)2016-03-07 19:38:01
体育科技(2016年2期)2016-02-28 17:06:07
中国麻业科学(2015年5期)2015-12-28 06:22:11
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
自然资源遥感(2014年3期)2014-02-27 11:56:41
