
苏南地区农业土地利用方式改变对土壤理化及生物学性质的影响
2019-11-11 01:57:14牛昱涵王心怡李江叶张维国陈金林梁永超
植物营养与肥料学报 2019年10期
牛昱涵,施 曼,王心怡,李江叶,张维国,陈金林*,高 岩*,梁永超
(1 南京林业大学南方现代林业协同创新中心,南京 210037;2 江苏省农业科学院农业资源与环境研究所,南京 210014;3 浙江大学环境与资源学院,杭州 310012)
苏南地区是我国粮食主产区之一[1],19世纪80年代,粮食种植面积高达171.8 × 104公顷[2]。近年来,该区域越来越多的农田被改为果园[3],至2010年,农田面积已减少了32.8%[4],同时,该地区水蜜桃产业迅速发展,现无锡地区水蜜桃栽培面积已超过2330公顷,并带动周边常州、张家港等地发展桃园近6660公顷[5]。土地利用方式的改变伴随着施肥和管理模式的变化[6]。苏南地区长期实行夏水稻-冬小麦轮作制,通过秸秆还田、施用速效化肥提高土壤肥力,年化肥施用量达903 kg/hm2[7],年固碳量达42.23 Tg[8]。而由稻麦轮作田改为桃园后,虽然改为有机肥 (猪粪、菜籽饼等) 与化肥配施的施肥方式,然而为了追求水蜜桃更高的经济效益,过量的施肥带来养分淋失而造成的面源污染问题极为严重。据统计,苏南地区桃园氮、磷素周年投入量分别达846 kg/hm2[9]、213 kg/hm2[10],远高于氮素 (100~200 kg/hm2)[11]和磷素 (20~100 kg/hm2)[12]的桃园适宜投入量。
两种土地利用方式下施肥种类、施肥量及管理方式的不同必然会影响土壤理化性状和生物学特性,研究这些变化对土壤和肥料养分的转化与利用的影响,可为土地合理规划、养分高效利用及农业可持续发展提供重要的理论基础。
1 材料与方法
1.1 采样点概况
采样点位于常州市和宜兴市境内 (31°20′~31°40′N,119°52′~120°08′E),属北亚热带季风性湿润气候,年均降雨量1180 mm,相对湿度70%,年均气温16.5℃,无霜期200~240 天,土壤质地均为粉质粘土,其中砂粒、粉粒和粘粒含量分别为13.9%、57.1%和28.2%,由相同母质发育而来。该区域在20年前均为稻麦轮作田。宜兴市东下田村、石房村和三迁村土壤为典型的水稻土,20年来一直实行稻麦轮作,常规管理为分别于作物幼苗期、分蘖期和拔节期施用尿素、复合肥以及尿素加复合肥,周年施氮量约为450~750 kg/hm2,周年施磷量约为83.6 kg/hm2;由稻麦轮作田改造的桃园,施肥改为有机肥(猪粪、菜籽饼等)、复合化肥和有机无机复混肥(29.9%、31.9%和38.2%),周年氮素投入量约为850 kg/hm2,周年磷素投入量约为252 kg/hm2,并于秋季果实采摘后,以菜籽饼、猪粪等有机肥为主施入基肥,追肥在桃树生长季进行2~3次,以化肥为主。上述6个采样点具体地理位置分布如图1所示。
1.2 土壤样品采集
土壤样品分别在5月 (麦季) 和7月 (稻季) 采集。在稻麦轮作田和桃园采样点,分别随机选择3个约为100 m × 100 m的地块进行采样,稻麦轮作田记为P1、P2和P3,桃园记为O1、O2和O3。将每个100 m × 100 m的地块等距划分为3个30 m × 30 m的小区作为3个重复,在每个小区按“S”型取样取9钻0—20 cm的土柱,置于干净塑料薄膜上,去除石砾、根系和动物残体等杂质,充分混匀,取约1 kg新鲜土样装入塑料密封袋,置于冰盒中供生物学性状测定,然后,用四分法取1 kg混合土样供化学性质测定。另外,每个样点采集3个环刀样品供土壤物理指标测定。冰盒中的新鲜土样带回实验室后,立即保存于 -20℃冰箱供生物学性状测定,混合土样置于阴凉通风处,经风干后,磨碎、过筛备用。
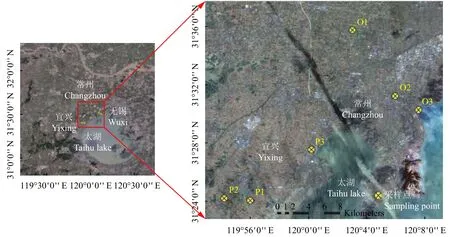
图1 采样点位置Fig. 1 Sampling location[注(Note):P1、P2和P3分别代表东夏田村、石房村和三迁村的稻麦轮作田;O1、O2 和 O3 分别代表后庄村、杭头上村、太湖头村桃园,已分别改为桃园4年、8年和20年。P1,P2 and P3 represent rice-wheat rotation fields in Dongxiatian,Shifang and Sanqian Village,respectively; O1,O2 and O3 represent peach orchards in Houzhuang,Hangtoushang andTaihutou Village,where the peach orchards has been operated for 4,8,and 20 years,respectively.]
1.3 试验方法
1.3.1 土壤物理性质测定 土壤容重和孔隙度采用环刀法测定[13]。土壤机械组成采用比重计法测定[14],并根据美国制土壤质地分类法确定土壤质地类型。土壤含水率采用烘干称重法测定[15],105℃连续烘干24小时后测定。
1.3.2 土壤化学性质测定 将风干土样过2 mm筛并混合均匀,然后部分过0.25 mm筛后测定其化学性质。土壤pH采用电位法测定 (土∶水 = 1∶2.5)[16]。有机碳采用重铬酸钾氧化—分光光度法测定[17]。总氮(TN) 采用凯氏定氮法测定[18]。铵态氮采用靛酚蓝比色法测定[15]。硝态氮采用紫外分光光度法测定[19]。全磷 (TP) 采用酸熔钼锑抗比色法测定[15]。有效磷 (AP)采用碳酸氢钠浸提—钼锑抗比色法测定[20]。
1.3.3 土壤生物学性质测定 1) 土壤酶活性测定采用96微孔酶标板荧光分析法测定[21],共测定了8种土壤酶活性,具体酶及其底物见表1及荧光定量PCR。
2) 土壤DNA的提取及荧光定量PCR 细菌16S和真菌ITS rRNA基因丰度的定量分析按照FastDNA Spin 试剂盒 (MP Biomedical,Carlsbad,USA) 说明书提取土壤DNA,其中最后一步改用80 Ml DES洗脱DNA。用Nanodrop 2000检测提取DNA的质量 (260/280 = 1.8~2.0) 并测定其浓度,将提取的土壤DNA样品保存于 -20℃冰箱中。
取上述DNA提取液5 μL,用灭菌水稀释10倍,用于细菌16S和真菌ITS rRNA基因丰度的定量测定。细菌16S rRNA和真菌ITS rRNA基因的荧光定量扩增的引物分别是515F/806R (515F,5′-GTGYCAGCMGCCGCGGTAA-3′;806R,5′-GGACTACNVGGGTWTCTAAT-3′) 和 ITS3/ITS4(ITS3,5′-GCATCGATGAAGAACGCAGC-3′;ITS4,5′-TCCTCCGCTTATTGATATGC-3′)。采用实时定量聚合酶链式反应 (RT-qPCR) 检测细菌和真菌的丰度反应,反应体系和程序参照Li等[22]的方法。反应在ABI7500 实时荧光定量系统 (Applied Biosystems,Santa Clara,CA) 上进行。细菌16S和真菌ITS rRNA基因丰度检测的标线使用的两个质粒的初始浓度分别对应的基因拷贝数为每克干土1.22 ×1010和9.05 × 1010copies。细菌和真菌荧光定量扩增的标线均使用10倍系列稀释的质粒作为模板,使用的数量级为108~102。其扩增效率分别为98%和99%,线性回归方程的R2分别为0.998和0.999。
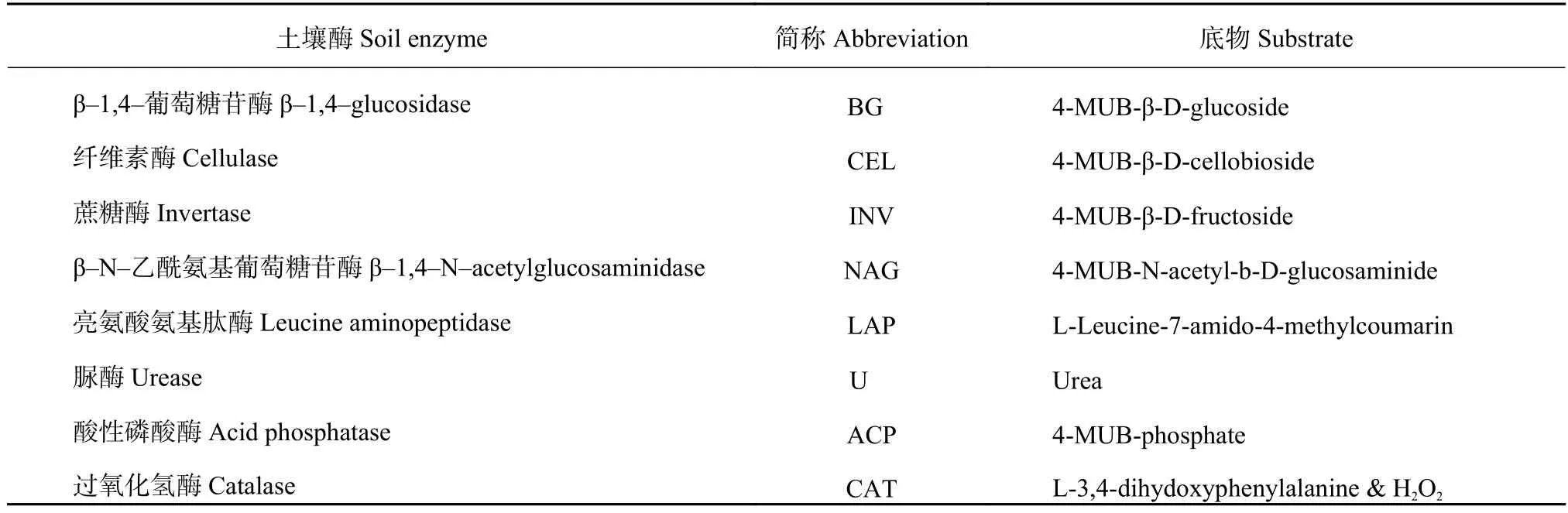
表1 检测的土壤酶及底物Table 1 Soil enzymes and substrates tested
1.4 数据分析
数据处理采用Excel 2007。SPSS 20.0用于单因素方差分析和皮尔逊相关分析,不同处理重复的平均值的统计差异采用Duncan检验,显著性水平为0.05或0.01。
作图软件为Sigmaplot 12.0。在对酶活性、细菌16S rRNA、真菌ITS rRNA基因丰度进行统计分析前先对测定的数据进行自然对数转换,以满足残差正态性和方差齐性的假设。由于不同改种年限(4年、8年和20年) 的桃园土壤理化性质和生物学特性之间的差异不显著 (P > 0.05),因此,将不同年限的处理合并为一种土地利用方式 (桃园) 处理。
2 结果与分析
2.1 不同利用方式对耕地土壤物理性状及pH的影响
在苏南地区,不论是传统的稻麦轮作田还是改种的桃园,土壤容重在5月和7月间差异均不显著(图2A),但同一季节,两种土地利用方式土壤容重的差异显著 (P < 0.05)。5月和7月,桃园土壤的容重均显著高于稻麦轮作田土壤,分别提高了9.0%和20.9% (P < 0.05)。
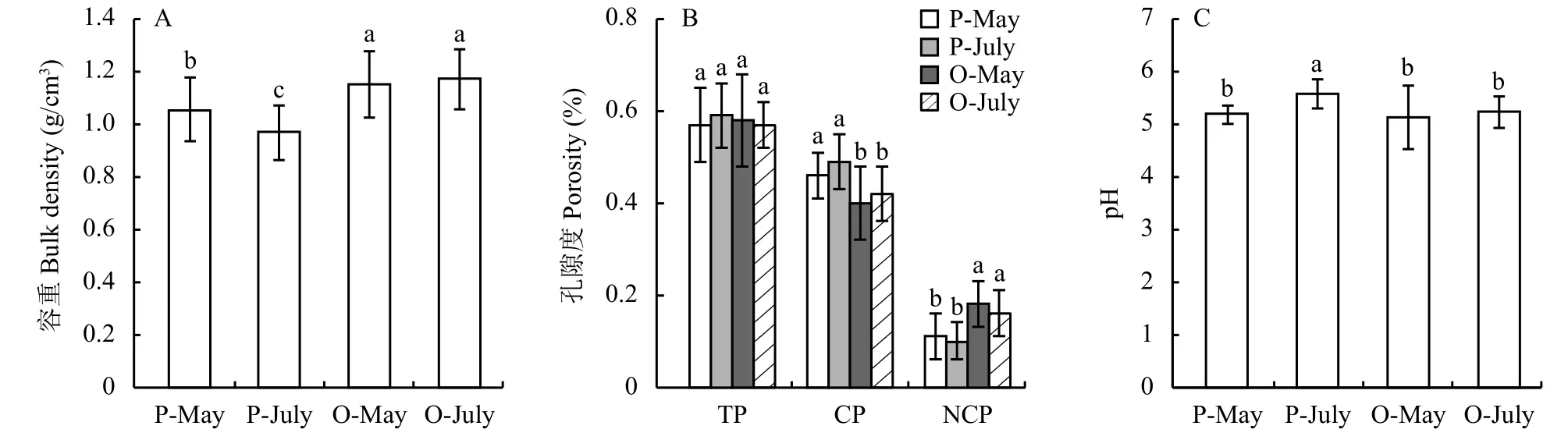
图2 不同土地利用方式下的土壤理化性质Fig. 2 Physical and chemical properties of soils under different land use types[注(Note):P-May—小麦生长季水稻土,于5月底采集Paddy soil collected in May during wheat season;P-July—水稻生长季水稻土,于7月底采集Paddy soil collected in July during rice season;O-May—桃园土,采于5月底Orchard soil collected in May;O-July—结果期桃园土,采于2017年7月底Orchard soil collected in July during peach fruiting stage;TP—总孔隙度Total porosity;CP—毛管孔隙度Capillary porosity;NCP—非毛管孔隙度Non-capillary porosity;柱上不同字母代表处理间差异达5%显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
不同土地利用方式下,土壤总孔隙度无显著性差异 (图2B),而土壤毛管孔隙度和非毛管孔隙度差异较大 (P < 0.05)。桃园的土壤毛管孔隙度比稻麦轮作田降低了13.8% (P < 0.05);相反,非毛管孔隙度比稻麦轮作田提高了57.7% (P < 0.05)。两种利用方式的毛管孔隙度都显著高于非毛管孔隙度 (P < 0.05)。
稻麦轮作田和改种后的桃园土壤的pH在4.08~6.42之间 (图2C),均值分别为5.39和5.19,桃园土壤pH显著低于稻麦轮作田 (P < 0.05)。在5月,麦田和桃园土壤pH无显著差异,而7月稻田土壤的pH明显升高,且显著高于桃园土壤和5月的麦田土壤 (P < 0.05)。
2.2 不同利用方式对耕地土壤有机碳和养分的影响
由表2可见,在5月,桃园土有机碳含量比稻麦轮作田低20.7% (P < 0.05),而7月无显著差异。桃园土有机碳含量在5月和7间无显著差异,而7月稻麦轮作田的土壤有机碳含量比5月降低了26.1% (P < 0.05)。
不论是在5月还是7月,稻麦轮作田土壤的全氮含量与桃园无显著差异;在5月,稻麦轮作田土壤的铵态氮和硝态氮分别比桃园土壤低75.0%和41.2% (P < 0.05),在7月,稻麦轮作田土壤的铵态氮含量无显著差异,而硝态氮含量比桃园土壤低71.4%(P < 0.05);在5月,桃园土壤全磷和有效磷含量分别是稻麦轮作田的3倍和6倍多。在7月,两者全磷含量无显著差异,桃园土壤有效磷含量约为稻田土壤的6.6倍。同种土地利用方式下,7月稻麦轮作田土壤全氮含量比5月下降了20.9%,桃园土壤下降了13.6% (P < 0.05);稻麦轮作田5月和7月土壤铵态氮含量差异不显著,7月硝态氮含量比5月下降了61.4% (P < 0.05)。7月桃园土壤铵态氮含量比5月下降了75.0% (P < 0.05),而硝态氮差异不显著。
2.3 不同土地利用方式对土壤生物学特性的影响
由图3可见,桃园土壤β-1,4-葡萄糖苷酶 (BG)活性在5月和7月分别比稻麦轮作田高4.7%和19.4%,纤维素酶 (CEL) 和蔗糖酶 (INV) 活性在相同季节均没有显著差异。在7月,稻麦轮作田的土壤CEL酶和INV活性比5月降低了12.6%和21.2%,桃园则分别降低了17.1%和25.0%。皮尔逊相关分析(表3) 表明,土壤BG、CEL、INV酶活性与有机碳含量呈显著正相关 (P < 0.05)。土壤BG酶活性还与硝态氮和有效磷含量呈极显著正相关 (P < 0.05),表明土壤碳转化相关的酶活性受到氮、磷养分的调控。
桃园土壤β-N-乙酰氨基葡萄糖苷酶 (NAG) 活性在5月和7月分别比稻麦轮作田低10.3%和15.3%(图3)。在7月份,稻麦轮作田和桃园土壤的脲酶活性分别比5月份下降了10.3%和9.7%,相关分析表明,土壤脲酶活性与土壤总孔隙度和毛管孔隙度呈极显著负相关 (表3,P < 0.01),相关系数分别为-0.349和-0.264。桃园与稻麦轮作田土壤的亮氨酸氨肽酶 (LAP) 活性在5月和7月都无显著性差异。另外,在7月份,桃园和稻麦轮作田土壤的NAG酶活性分别比5月降低了31.3%和27.2%,相关分析表明土壤NAG酶活性与土壤有机碳和全氮含量呈极显著正相关 (表3,P < 0.01),表明土壤NAG酶活性除受到氮元素限制之外,还可能受到碳元素的调控。
在7月份,桃园土壤的酸性磷酸酶 (ACP) 活性比稻麦轮作田低8.5% (图3)。相同土地利用方式下,7月桃园和稻麦轮作田的ACP酶活性分别比5月下降了21.1%和14.0%,相关分析表明土壤ACP酶活性与土壤有机碳、全氮、全磷含量呈极显著正相关 (表3,P < 0.01),而与有效磷无显著相关性。表明桃园和稻麦轮作田土壤与磷转化相关的酶活性除受磷元素限制外,还可能受到碳、氮元素的调控。
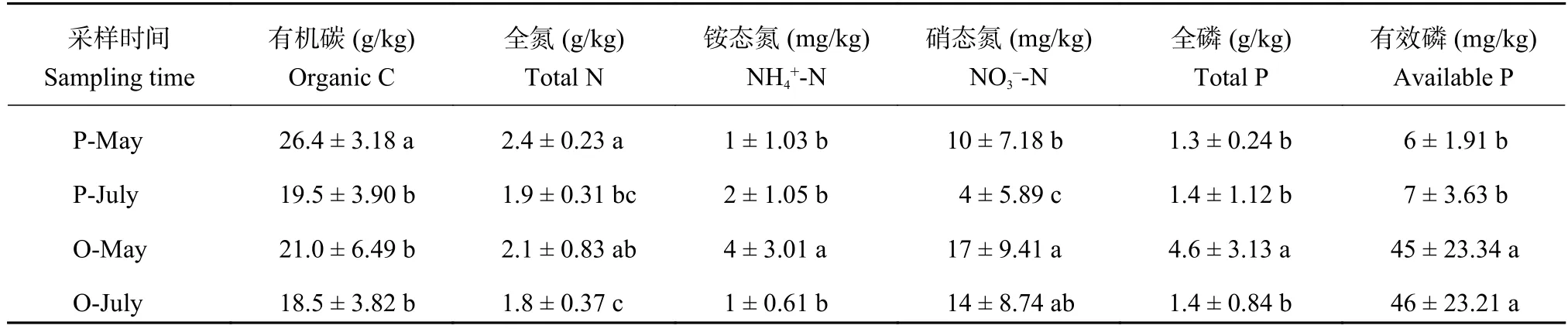
表2 不同土地利用方式下的土壤化学性质Table 2 Soil chemical properties under different land use types
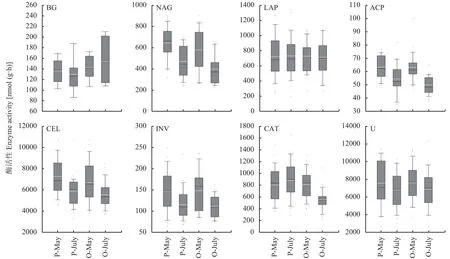
图3 不同土地利用方式下土壤酶活性Fig. 3 Soil enzymatic activities under different land use types[注(Note):P-May—小麦生长季水稻土,于5月底采集Paddy soil collected in May during wheat season;P-July—水稻生长季水稻土,于7月底采集Paddy soil collected in July during rice season;O-May—桃园土,采于5月底Orchard soil collected in May;O-July—结果期桃园土,箱式图中的虚线代表数据的平均值,实线代表数据的中位数The dotted line in the box plot represents the average of the data,and the solid line represents the median of the data.]
在5月份,桃园和稻麦轮作田的土壤过氧化氢酶 (CAT) 活性没有显著性差异 (图3),在7月份,桃园的土壤CAT酶活性比稻麦轮作田低37.3%。相关分析表明土壤CAT酶活性与土壤含水率、有机碳、全氮、铵态氮都呈显著正相关 (表3,P < 0.05),表明土壤CAT酶可能同时受土壤水分和碳氮元素的调控。
桃园土壤的细菌丰度在5月和7月分别比稻麦轮作田低55.1%和44.1% (图4),桃园土壤的真菌丰度在5月和7月分别比稻麦轮作田高176%和310%。桃园和稻麦轮作田的细菌丰度在7月份比5月份分别下降了25.1%和39.8%,真菌丰度分别降低了39.2%和59.0%。皮尔逊相关分析表明,土壤细菌16S rRNA基因丰度与土壤pH、含水率呈显著正相关 (P < 0.05),与有效磷含量呈极显著负相关 (P <0.01);土壤真菌ITS rRNA基因丰度则相反,与土壤pH、含水率呈极显著负相关 (P < 0.01),与有效磷含量呈极显著正相关 (P < 0.01)。这些结果表明土壤微生物群落在细菌和真菌占比的变化受pH、含水率、有效磷含量的影响。此外,土壤细菌16S rRNA基因丰度与NAG、ACP和CAT酶活性呈显著正相关 (P < 0.05),而真菌ITS rRNA基因丰度则与各土壤酶活性相关不显著,表明土壤细菌可能对土壤氮、磷养分转化影响较大,同时可能影响过氧化氢等物质的降解。
3 讨论
3.1 不同利用方式对土壤性状的影响
3.1.1 不同利用方式对土壤物理性状的影响 总体上,稻麦轮作农田和改种为桃园的土壤容重在1.1~1.4 g/cm3之间,总孔隙度均高于55%,且土壤孔隙结构以毛管孔隙为主 (图2B),而非毛管孔隙度超过10%,均保持着良好的物理结构[23]。但由于稻麦轮作田和桃园实行不同的耕作、施肥等管理措施,两种利用方式下土壤容重、含水率、孔隙度等物理性质仍呈现一定差异。稻麦轮作田的土壤容重较低,这可能与稻麦轮作农田实行长期的机械深松和秸秆还田有关:一方面,经过深松的土壤疏松、孔隙增大、容重减小;另一方面,秸秆还田也增加了土壤孔隙度,从而降低土壤容重[24]。较小的土壤容重意味着土壤团聚体稳定性较高,土壤物理结构良好,土壤自然含水率高,田间持水能力强,土壤水、气、热条件适宜[25]。桃园的土壤容重较高,主要是因为桃树的生长周期较长,长期不翻耕和踩踏使土壤更加紧实。此外,稻麦轮作田改为桃园后,土壤孔隙由毛管孔隙向非毛管孔隙转化,表明稻麦轮作田改种为桃园后土壤的持水性和透气性能变差。
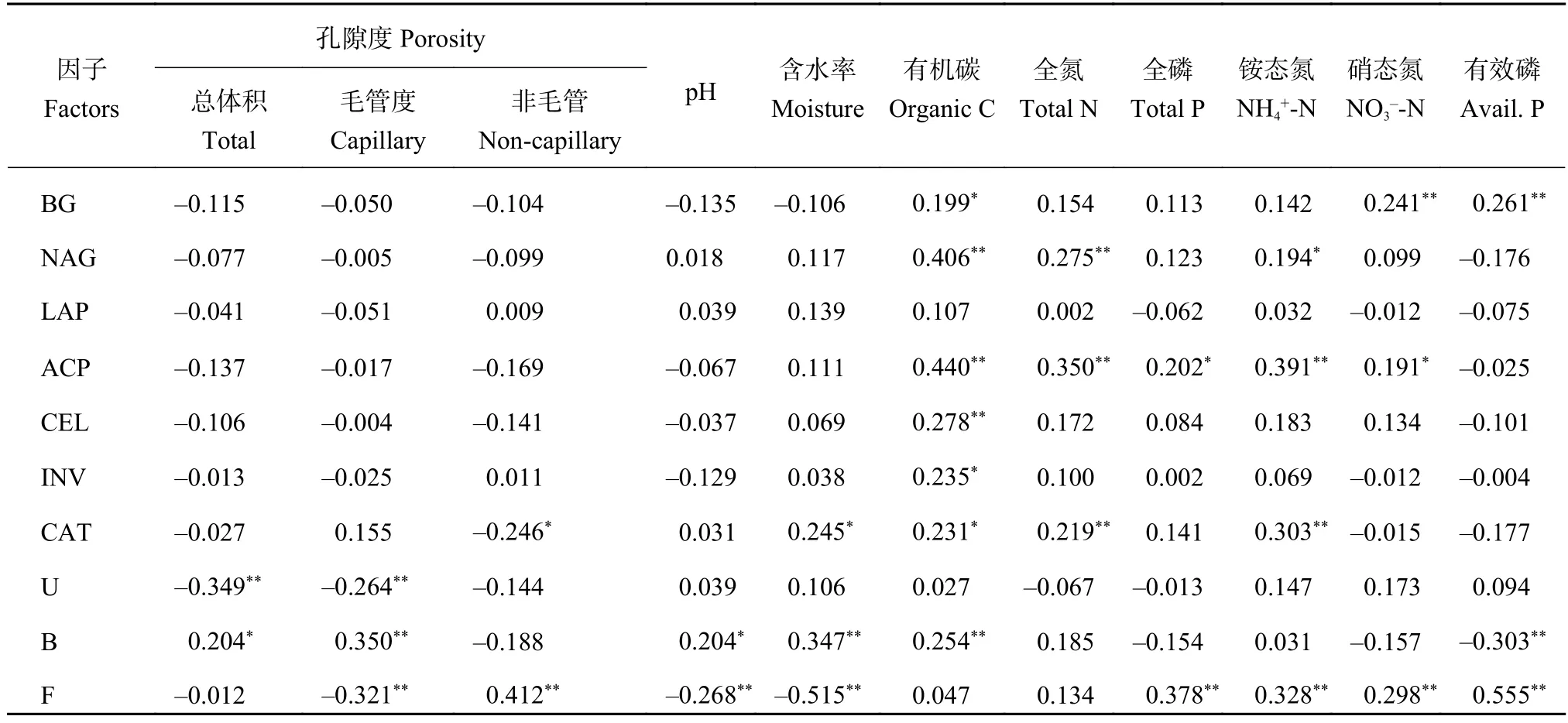
表3 土壤生物特性和与土壤理化因子的皮尔逊相关分析Table 3 Pearson product-moment correlation analysis of the relationships between soil biological properties and soil physiochemical factors
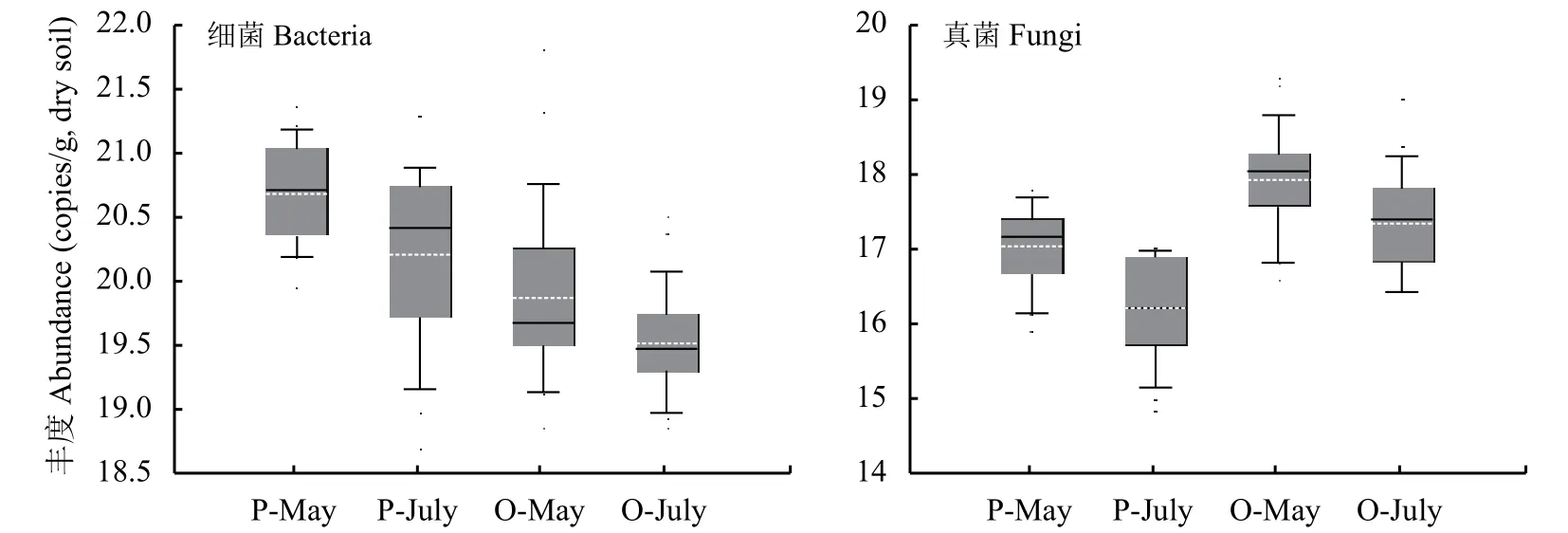
图4 不同土地利用方式下的土壤细菌16S rRNA基因和真菌ITS rRNA基因丰度Fig. 4 Abundance of bacterial 16S rRNA genes and fungal ITS rRNA genes in soils under different land use types[注(Note):P-May—小麦生长季水稻土,于5月底采集Paddy soil collected in May;P-July—水稻生长季水稻土,于7月底采集Paddy soil collected in July;O-May—桃园土,采于5月底Orchard soil collected in May;O-July—结果期桃园土,采于2017年7月底Orchard soil collected in July;箱式图中的虚线代表数据的平均值,实线代表数据的中位数The dotted line in the box plot represents the average of the data,and the solid line represents the median of the data.]
3.1.2 不同利用方式对土壤化学性状的影响 稻麦轮作田和桃园土壤的pH在4.08~6.42之间,平均值分别为5.39和5.19,较2000年分别下降10.3%和13.7%,与1979年相比分别下降15.4%和18.5%[26]。稻麦轮作田和桃园土壤酸化加剧的原因一方面是由于自然因素,苏南地区的土壤主要为黄泥土棕壤,属于弱酸性母质,自然发育易形成酸性物质,加之土壤碱性盐基离子淋溶使土壤上层盐基饱和度下降引起土壤酸化[27];另一方面因为两种利用方式均长期施用大量复合肥、尿素等含氮肥料,NH4+的硝化作用会产生大量H+导致土壤酸化[28]。尤其是桃园的施氮量[9]远高于桃树需求量[11],因此土壤呈现酸化趋势,过量的施肥也是桃园土壤pH显著低于稻麦轮作田的主要原因 (P < 0.05)。此外,稻麦轮作田干湿交替后土壤pH出现明显变化,淹水条件下土壤pH升高的原因一方面可能是由于土壤反硝化反应消耗H+[29],另一方面可能与土壤中的铁、锰和硫的还原反应消耗H+有关[30]。土壤酸化会造成一系列生态环境和农产品安全问题[31]。因此,苏南地区在经济利益趋动下,土地利用方式由稻麦轮作改为桃园导致的土壤酸化问题应引起关注。
本研究结果显示,稻麦轮作田和桃园土壤的有机碳、全氮、硝态氮、全磷、有效磷含量都高于同区域的森林土壤[32],这是由于两种土地利用方式下氮磷及有机物料的投入量过高导致的。两种土地利用方式相比,稻麦轮作田土壤的有机碳含量高于桃园,这主要是由于稻麦轮作长期实施秸秆还田,向土壤输入了大量碳氮比较高的有机物料,使得稻田土壤有机碳得到长期累积,而桃园施用的有机肥种类主要是鸡粪、猪粪等碳氮比低的有机物料,容易被微生物分解利用,不利于土壤有机碳的累积[27]。
稻麦轮作田和桃园土壤中的硝态氮含量远高于铵态氮的含量,这可能是施入土壤的铵态氮通过氨挥发和土壤硝化作用的结果。研究表明农田施用的铵态氮中约11%通过氨挥发损失[33],60%~80%转变成硝态氮[34]。两种土地利用方式相比,桃园土壤的硝态氮含量高于稻麦轮作田,且5月份桃园土壤的铵态氮含量也高于稻麦农田,这是由于两种土地利用方式施用的氮肥均以铵态氮肥为主,而桃园以农家肥作为基肥,有文献报道,农家肥中的铵态氮含量高于化肥[35],因此,与7月份相比,5月份气温较低,桃树所处生长期对土壤肥料的需求也较低,土壤中的铵态氮盈余,这也为桃园土壤中高浓度的硝态氮提供了来源。另外,两种土地利用方式土壤硝态氮含量也存在较大差异,其原因一方面是稻麦轮作田在淹水时期,嫌气条件下强烈的反硝化作用使土壤硝态氮严重损失[36],另一方面水田土壤中的硝态氮也可能通过淋洗和田面径流损失[37]。
桃园土壤的全磷和有效磷含量均显著高于稻麦轮作田,这主要与施肥量和种类有关。桃园磷肥的年均纯养分用量为252 kg/hm2,约为水田年均磷肥用量的3倍[10]。稻麦轮作田主要施化肥,而桃园主要施用粪肥、豆饼等有机肥,其中磷的含量远高于化肥且易释放[38]。
3.1.3 不同利用方式对土壤生物学性状的影响 土壤微生物会在环境变化时通过改变分泌和释放土壤酶来适应环境[39],因此土壤理化性质的改变会影响微生物对养分的获取,从而影响土壤酶的分泌。本研究中,土地利用方式由稻麦轮作田转变为桃园后,土壤生物学性质发生显著变化,表现为较稻麦轮作田,桃园的BG酶活性升高,NAG、U、ACP、CAT酶活性降低。这可能是稻麦轮作田采用含大量有机物质的秸秆和无机肥配合施用,可以增强土壤微生物代谢活性,提高多种水解酶的活性,从而有效促进秸秆矿化和养分释放。同时,嫌气条件下分解的有机质矿化容易产生大量过氧化物,对微生物生长不利,微生物为消除其毒害影响,会分泌大量过氧化物酶[40],导致稻麦轮作田CAT酶活性较高。
从微生物群落来看,与稻麦轮作田相比,桃园的土壤细菌16S rRNA基因丰度下降,真菌ITS rRNA基因丰度升高,这可能与以下三个方面有关:一是桃园土壤pH较稻麦轮作田可能呈现出较大的降低趋势,绝大多数真菌适宜在酸性环境中生长,而细菌主要适宜在中性环境[41-43],导致土壤由“细菌型”向“真菌型”转变[41]。二是桃园的土壤有效磷含量明显高于稻麦轮作田,磷素施入会显著提高土壤真菌数量和真菌/细菌比值[43-45],导致土壤微生物群落由细菌向真菌转化。三是桃园土壤的含水率较低,在较干旱的环境下,由于真菌自身的特殊生理结构,如菌丝和孢子,使得真菌的生存能力强于细菌[46],造成水分状况较差的桃园土壤微生物群落中真菌ITS rRNA基因丰度较高。
3.2 不同季节对土壤性状的影响
从季节角度来看,稻麦轮作田土壤的有机碳含量整体高于桃园。但从5月到7月也就是从春季过渡到夏季,稻麦轮作田有机碳含量的下降幅度较大。这一方面可能是由于稻田淹水导致土壤有机碳矿化速率加快,如研究表明,随土壤持水量升高而增加[47-49]。另一方面,这可能与稻麦轮作田夏季温度较高有关。随着表层土壤温度上升微生物活性增强[50],导致有机碳矿化比例增加[51-53],而桃树的遮阴作用使土壤有效避免阳光直射,平均土壤温度比稻麦农田低5.3%,因此有机碳矿化速率低于稻麦轮作田[54]。
在春节和夏季,稻麦农田土壤铵氮浓度无显著差异 (P < 0.05),但夏季土壤硝态氮浓度却显著低于春季土壤 (P < 0.05),一方面可能是由于干湿交替导致土壤淹水进入嫌气状态,硝态氮的淋溶和反硝化作用增强[55-57],另一方面温度升高导致植物光合作用增强和根系生长,产生更多的根际分泌物和沉积物[58],为反硝化微生物提供更多的能源,促进微生物生长[59]。同时,适度升温可促进土壤反硝化速率[60]。桃园夏季土壤中硝态氮的减少主要是由于土壤铵态氮的降低,桃园施用的氮肥主要是尿素、农家肥等铵态氮含量较高的肥料[33],土壤中的硝态氮主要是由铵态氮硝化而来,而在夏季桃树结果期需要大量吸收速效氮,导致土壤中铵态氮含量大幅下降,硝态氮含量因来源减少也随之下降。
稻麦轮作田和桃园的土壤细菌16S rRNA基因丰度和真菌ITS rRNA基因丰度在夏季较春季都显著下降。这一方面是源于土壤养分变化[61],土壤的有机碳、全氮和硝态氮含量均在夏季显著降低,因作物和桃树生长旺盛,与土壤微生物竞争有效养分,微生物的矿化作用大于固持作用[62],细菌和真菌可利用的碳和养分减少导致其丰度下降,这也可以从春季过渡到夏季土壤中纤维素酶和蔗糖酶的活性下降反映出来 (图3)。另一方面是由于在稻麦轮作田,稻季长时间淹水导致土壤处于缺氧状态,大部分土壤微生物属于好氧型微生物[63]。因此,水淹作为胁迫因子可能导致土壤微生物丰度下降[64]。桃园微生物丰度下降的主要原因可能与结果期的管理措施有关,7月是桃果成熟、收获的季节,农民对桃园的踩踏频繁,使得土壤变紧实、通气性变差,导致微生物的生境条件变差、丰度下降。
4 结论
在苏南地区,随着土地利用方式由稻麦轮作田转变为桃园,土壤理化、生物学性质发生明显变化,土地施肥、管理措施是导致土壤性质变化的主要原因。桃园施用农家肥更多,翻耕减少,人为管理更加频繁。土地利用方式变化首先影响土壤物理性质,使土壤更加紧实;其次影响土壤化学性质和养分状况,使土壤pH有进一步降低的趋势,土壤有机质含量下降,全磷和有效磷含量增加;进而影响土壤微生物丰度和酶活性,土壤细菌16S rRNA基因丰度下降,真菌ITS rRNA基因丰度升高,有机质降解的微生物群落可能由“细菌型”向“真菌型”转化。相应地,土壤中与氮转化相关酶和过氧化氢酶活性降低。
从春季到夏季,因温度升高、稻田淹水、桃园管理等原因,稻麦轮作田和桃园土壤有机碳含量均下降,稻田降幅较大。相应地,两种利用方式下土壤的细菌16S rRNA基因丰度、真菌ITS rRNA基因丰度、纤维素酶和蔗糖酶的活性均显著下降。
猜你喜欢
幼儿100(2023年21期)2023-06-21 02:20:46
社会科学战线(2022年1期)2022-02-16 01:10:26
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29 03:15:14
农民致富之友(2020年31期)2020-11-18 08:49:34
农业环境科学学报(2020年10期)2020-11-11 02:55:28
河北果树(2020年2期)2020-05-25 06:58:50
民族音乐(2019年3期)2019-08-14 01:05:18
小资CHIC!ELEGANCE(2018年8期)2018-04-03 05:55:46
安徽农学通报(2017年11期)2017-06-23 18:34:49
浙江农业学报(2016年7期)2016-06-15 20:30:11
