
农田土壤N2O排放的关键过程及影响因素
2019-11-11 01:57:28曹文超王娅静郭景恒王敬国
植物营养与肥料学报 2019年10期
曹文超,宋 贺,王娅静,覃 伟,郭景恒,陈 清,王敬国*
(1 中国农业大学资源与环境学院,北京 100193;2 中国热带农业科学院环境与植物保护研究所,海南海口 571101;3 安徽农业大学农学院,安徽合肥 230036;4 河北农业大学资源与环境科学学院,河北保定 071001)
一氧化二氮 (N2O,俗称氧化亚氮) 作为重要的温室气体之一,其全球增温潜势为CO2的190~270倍,且浓度以每年0.25%的速率持续增加,加剧了全球气候变暖[1]。同时,N2O也是破坏臭氧层的主要因子[2]。因而,N2O的排放在全球气候和生态环境变化的研究中受到广泛关注。
农业活动是N2O最大的人为排放源,其中农田土壤对N2O排放贡献最大[3-5]。农田土壤每年N2O排放约占全球人为活动N2O排放总量 (6.7 × 106t) 的42%[6]。以我国农田土壤为例,2014年N2O排放量约为1.21 Tg,占全球N2O排放量的31%[7]。一般认为农田土壤N2O产生的主要途径为硝化和反硝化作用过程[8],同时,环境因子 (土壤pH、温度等) 和管理措施 (施肥、灌水等) 主要通过影响这两个过程进而影响N2O排放。然而,综合土壤N2O排放的其他过程并结合相关环境因子的分析仍缺乏系统性研究。因此,探究农田土壤各调控机制和环境因子对N2O排放的影响,有利于理解农田土壤氮素转化过程与N2O排放之间的关系,从而为进一步提出合理的N2O减排措施提供理论依据。
1 农田土壤产生N2O的过程
农田土壤产生N2O的生物学过程包括硝化作用、反硝化作用、硝化细菌的反硝化作用、硝酸异化还原成铵以及真菌反硝化作用等。通常认为反硝化和硝化作用是产生N2O的主导途径。近年来在土壤低氧环境条件下硝化细菌的反硝化作用对N2O排放的贡献受到重视。此外,硝化-反硝化耦合作用和真菌反硝化作用对N2O排放也有贡献。而且,化学反硝化、亚硝基化反应 (nitrosation) 等非生物学过程也是土壤N2O的产生途径 (图1)。因此,全面了解与N2O排放有关的土壤过程,有助于实现农田土壤N2O减排。
1.1 硝化作用
硝化作用 (nitrification) 通常有两个过程:氨氧化过程和亚硝酸氧化过程 (图1A)。氨氧化过程是自养的氨氧化细菌 (ammonia-oxidizing bacteria,AOB) 或氨氧化古菌 (ammonia-oxidizing archaea,AOA) 在氨单加氧酶 (AMO) 催化下将NH3催化氧化为羟胺(NH2OH),并进一步在羟胺氧化还原酶 (HAO) 催化下产生NO2-,该过程是硝化作用发生的限速步骤[9]。亚硝酸氧化是指NO2-在亚硝酸氧化菌 (nitrite oxidation bacteria,NOB) 作用下氧化为NO3-的过程。最近研究发现了完全氨氧化菌 (如Nitrospira) 的存在,这类细菌能够将铵态氮直接氧化为硝态氮,称为完全硝化过程 (complete ammonia oxidizers,comammox)[10-11],这一发现颠覆了硝化过程必须经过两步反应过程的经典认知。除自养硝化作用外,异养硝化作用是指异养微生物将有机态氮氧化为亚硝态氮或硝态氮的过程,其中参与该活动的微生物有细菌、真菌等[12-13]。
N2O不是硝化过程的中间产物,其产生与中间产物NH2OH和NO2-紧密相关。硝化作用对N2O排放的直接贡献较低,大多低于NH4+-N氧化总量的1%[14]。纯培养发现,不同亚硝化螺菌氨氧化过程产生的N2O与其累积的NO2-比例为 0.07%~0.1%[15]。Mørkved等通过泥浆培养试验发现,土壤硝化作用产生的N2O与硝化终产物NO3-的比值也仅为0.02%~0.09%[16]。硝化过程对土壤N2O排放的贡献,或许主要体现在其为包括经典生物反硝化过程、硝化细菌的反硝化过程,甚至非生物的N2O产生过程等提供NO3-和NO2-等底物上,其中氨氧化过程产生的终产物NO2-尤为重要。
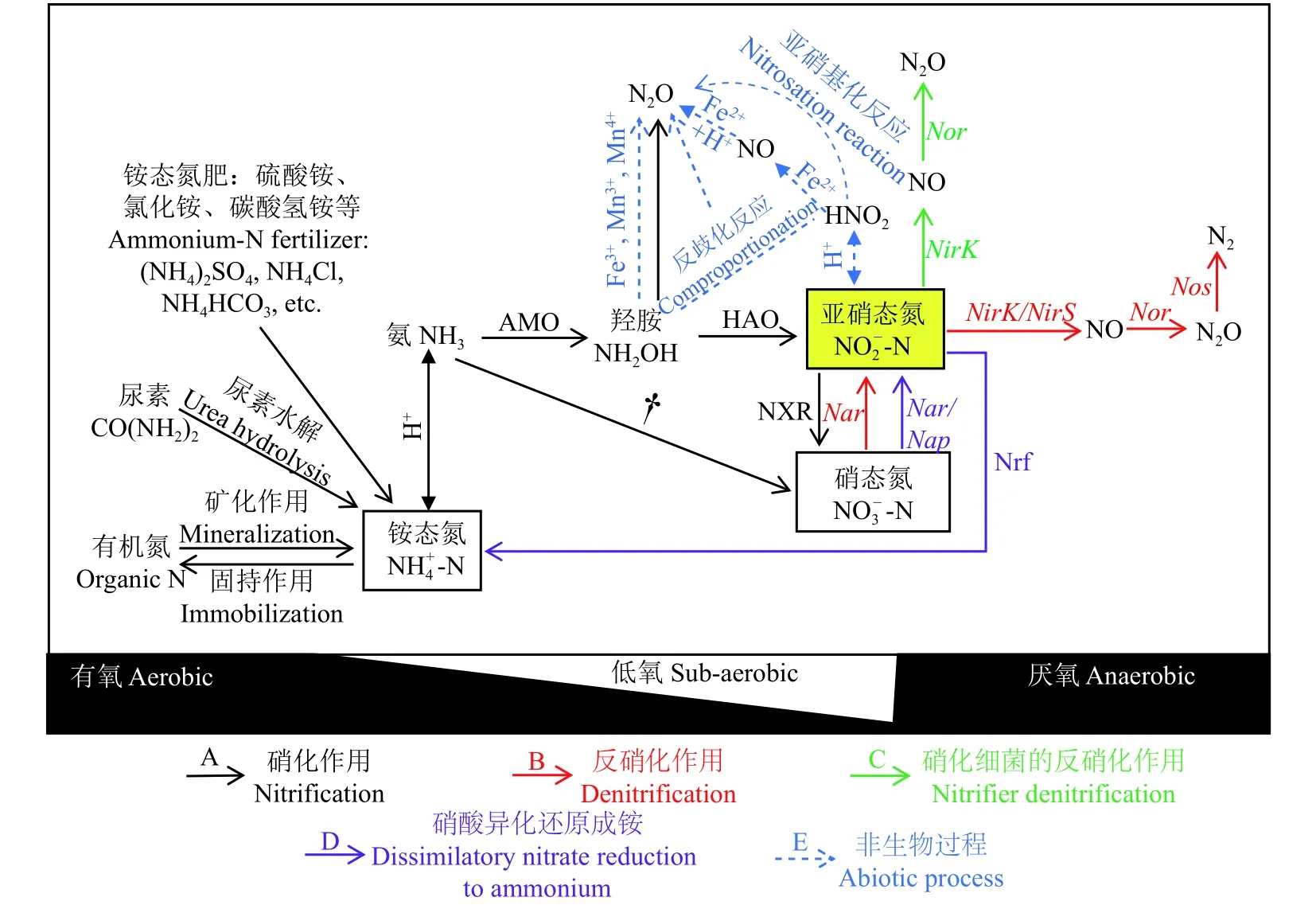
图1 N2O排放的主要生物学和非生物学过程Fig. 1 Biotic and abiotic processes of nitrous oxide (N2O) emission from soils[注(Note):AMO—氨单加氧酶Ammonia monooxygenase;HAO—羟胺氧化还原酶Hydroxylamine oxidoreductase;NXR—亚硝酸氧化还原酶Nitrite oxidoreductase;Nar/Nap—硝酸异化还原酶Membrane-bound nitrate reductase/periplasmic nitrate reductase;Nrf—亚硝酸还原酶Cytochrome c nitrite reductase;NirK—含铜的亚硝酸异化还原酶Copper-containing nitrite reductase;NirS—含有细胞色素cd1的亚硝酸异化还原酶Cytochrome cd1 nitrite reductase;Nor—NO还原酶Nitric oxide reductase;Nos—N2O还原酶Nitrous oxide reductase;† 该过程 (一步硝化反应) 由Nitrospira细菌作用† This process (one-step nitrification) attributed only to the Nitrospira bacteria[10-11];A、B、C、D、E分别代表硝化、反硝化、硝化细菌反硝化、硝酸异化还原成铵及非生物学过程 A,B,C,D,E represent nitrification,denitrification,nitrifier deitrification,dissimilatory nitrate reduction to ammonium and abiotic processes,respectively.]
进一步对比自养硝化和异养硝化作用对N2O排放的相对贡献来看,一般认为异养硝化作用对N2O的贡献较少,但在某些土壤环境条件下不容忽视[17]。如,在酸性森林土壤中异养硝化作用是产生N2O的主要途径[18-23],其贡献主要受土壤pH和C/N影响[20]。此外,目前还暂未有证据显示,完全硝化过程 (comammox) 具有产生N2O的能力,但由于其生物化学过程是经典氨氧化细菌和亚硝酸氧化细菌相关过程的结合,其存在具有产生N2O的可能性[21]。
1.2 反硝化作用
反硝化作用 (denitrification) 通常是指生物反硝化,即反硝化微生物在厌氧条件下将NO3-或NO2-逐渐还原为NO、N2O和N2的过程 (图1B)[22]。该过程所需的酶包括异化硝酸盐还原酶 (Nar & Nap)、异化亚硝酸盐还原酶 (Nir)、一氧化氮还原酶 (Nor) 以及一氧化二氮还原酶 (Nos),相应的编码基因分别为narG和napA、nirK/S、norB以及nosZ。其中,NO2-还原为NO的过程被认为是反硝化作用中最关键的步骤[23]。亚硝酸盐还原酶 (Nir) 是产生NO的关键酶,包含由nirS基因编码的细胞色素还原酶 (cd1-Nir) 和由nirK基因编码的可溶性含铜酶 (Cu-Nir)[24]。反硝化微生物通常只含有其中的一种,已分离的反硝化细菌中含nirS的数量占3/4[25],而且在许多环境中以含有nirS基因的反硝化微生物占主导[25]。
通常反硝化作用是土壤产生N2O的主要来源。有研究表明,反硝化作用对N2O排放的贡献在农田、茶园和竹园土壤中分别为54.5%、72.8%和77.1%[20]。完全的反硝化作用终产物为分子态氮(N2),然而约有1/3数量的反硝化细菌缺少编码N2O还原酶 (N2OR) 的nosZ基因,导致其终产物为N2O[26]。同时,低土壤pH或高NO3-含量会抑制N2O还原酶的形成[27]。此外,在土壤碳有效性较低条件下,与其他异化还原酶相比,N2O还原酶竞争电子的能力较弱,因而也会造成N2O排放量增加[28]。
1.3 硝化细菌反硝化
硝化细菌的反硝化作用 (nitrifier denitrification,ND) 是氨氧化微生物直接利用其在氨氧化过程产生的亚硝酸盐 (NO2-) 作为电子受体,进行反硝化的过程。通常多发生在好氧或低氧条件下,进行该反应的微生物通常为氨氧化细菌 (AOB)[29],还未发现氨氧化古菌 (AOA) 具有这种还原能力[30]。此外,由于AOB缺乏N2OR编码基因,因而其终产物绝大部分是N2O,而非N2[31]。硝化细菌反硝化作用对土壤N2O排放可能占有较高比重[32-33]。Zhu等利用双同位素标记法证实在低氧浓度条件下,施用铵态氮肥后硝化细菌反硝化作用是土壤N2O排放的主要来源[34]。Huang等研究发现,集约化石灰性低碳旱作土壤中硝化细菌的反硝化作用占N2O排放总量的44%~58%[35]。但也有研究对此持怀疑态度[36],主要是由于定量硝化细菌反硝化的双同位素方法是基于氨氧化过程产生的NO2-只能被自养AOB还原的假设。然而,事实上许多异养反硝化菌也能够直接异化还原NO2-,从而导致高估了硝化细菌反硝化作用对N2O排放的贡献量。
1.4 硝化-反硝化耦合作用
一般认为,硝化和反硝化作用分别发生在有氧和厌氧条件下,即两个过程在不同的生态位中进行。硝化-反硝化耦合作用 (nitrification-coupled denitrification,NCD) 的实质是同时存在于两个相邻微域空间的硝化和反硝化过程的偶联;发生在有氧空间、由硝化过程产生的NO2-或NO3-作为底物直接被存在于厌氧或低氧空间的反硝化微生物所利用[12,37],从而导致在好氧-厌氧界面上N2O的排放。
硝化-反硝化耦合作用对污水中的氮素去除、沉积物及农田土壤的氮素转化发挥着重要作用[37-38]。有氧条件的反硝化作用和沉积物内孔隙水的对流运输可使这两个过程在空间和时间上耦合更加紧密[39]。同时,研究已经证实,沉积物中添加15NH4+后,由反硝化作用产生的15N-N2几乎与氨氧化过程和亚硝酸氧化过程同步,即硝化和反硝化过程在发生时间上重叠[41]。在某些情况下硝化-反硝化耦合作用能够去除河口沉积物中10%~80%的人为氮素污染[40]。在土壤好氧微域环境中,如土壤表面氧化层或根际,硝化产生的NO3-或NO2-进入土壤厌氧微域会进一步发生反硝化作用。已有研究表明,在稻田土壤存在硝化-反硝化耦合作用,且硝化-反硝化耦合作用产生的氮损失很可能受硝化速率调控[41]。同时Ma等[42]研究发现,在稻田土壤250%和100%WHC条件下硝化-反硝化耦合作用对N2O排放总量的贡献比例分别为6.74%和11.5%。
1.5 真菌反硝化
真菌反硝化作用 (fungal denitrification) 主要是以NO2-作为底物在各种酶参与下还原为N2O的过程[43],其中某些真菌 (如Fusarium oxysporum) 也能够将NO3-还原为N2O。与细菌反硝化过程不同,由于真菌在线粒体膜上缺少N2OR,故不能将N2O进一步还原为N2[45]。进行反硝化作用的细菌和真菌均能利用NirK编码的亚硝酸盐还原酶将NO2-还原为NO[22,44]。与细菌不同的是,真菌主要是在NO还原酶细胞色素P450nor的作用下,以烟酰胺腺嘌呤二核苷(NADH,一种辅酶) 作为电子供体将NO还原为N2O[45],这种NO还原酶在结构和功能上与细菌NO还原酶有较大差别[46]。
一般认为,土壤N2O的产生以细菌主导的反硝化作用过程为主[47]。近年来,越来越多的证据显示,真菌在不同生态系统中对土壤N2O的排放均有贡献[48-52],尤其在酸性土壤条件下更明显[53-54,61],如真菌反硝化作用在低pH茶园土壤中对N2O排放的贡献高达70%[55]。这主要与真菌群落在低pH条件下占主导,而细菌对酸性土壤环境条件较为敏感有关[56]。相反,真菌反硝化作用在碱性土壤中对N2O的贡献并不重要[57]。在沙漠和半干旱的草地土壤中真菌也是N2O产生的主要来源[52],同时,在极端高温条件(40℃) 出现时,酸性土壤施用有机肥后可能也会增加真菌反硝化作用对N2O排放的贡献[58]。Wei等发现农田土壤田间表层施用颗粒性有机肥后N2O排放主要来源于真菌的反硝化作用[52]。
1.6 硝酸异化还原成铵
硝酸异化还原成铵 (dissimilatory nitrate reduction to ammonium,DNRA) 是硝酸异化还原的过程之一。与反硝化作用类似,DNRA过程是以NO3-或NO2-作为电子受体并在厌氧条件下将其还原为NH4+的过程 (图1D)。土壤pH、碳源含量、底物浓度(NO3-和NO2-)、C/N等是影响DNRA和反硝化作用这两个氮异化还原过程竞争的重要环境因素[59]。一般认为,DNRA更易在pH较高的条件下发生。如Yoon等已证实在酸性条件下Shewanella loihica PV-4菌株主要产物为N2O,而在碱性土壤中为NH4+[60]。同时,在NO3-或NO2-底物浓度较高或电子受体充足、碳源含量相对较低的环境中,由于DNRA过程发生所需的自由能比反硝化作用过程自由能高,因此该条件下反硝化作用更易发生[61]。相反,在电子受体 (NO3-/NO2-) 不足而碳源含量充裕时,DNRA过程则占主导[61]。
通常,DNRA过程在海洋生态系统的氮素循环中发挥重要作用,而陆地生态系统其多发生在湿地、草地等自然土壤中[62],其对N2O排放的贡献较低,甚至可以忽略。如Kool等通过15N-NH4+和15NNO3-标记试验表明,在沙壤土中DNRA过程对N2O的贡献可以忽略[63]。然而,也有研究发现具有DNRA功能的细菌中含有非典型N2OR,可将N2O进一步还原,这对N2O消耗具有潜在贡献[64]。
1.7 非生物学过程
除生物学过程外,近年来非生物学过程对N2O排放的贡献再次受到关注[65-68]。非生物学产生N2O的过程又称为化学反硝化作用 (chemodenitrification)[69],包括NO2-的分解、NO2-和NH2OH与金属离子等作用发生的电子转移过程[70]以及光化学反应等[71-72]。非生物学过程的发生主要与硝化、反硝化过程释放的不稳定中间产物NH2OH和NO2-有关 (图 1E)。它们可与金属离子 (如Fe2+或Mn3+) 或有机质发生非生物的化学反应产生N2O、NO,甚至N2[70]。其中,NO2-与有机物质的反应,被称为亚硝基化作用 (nitrosation)。一般而言,NH2OH很少在土壤中出现累积,这与NH2OH具有很高的活性且其氧化的发生与土壤pH、Mn和土壤有机质 (SOM) 的含量有关[73]。如Heil等研究发现,尽管土壤中Mn的含量较低,但NH2OH会优先与Mn3+或Mn4+进行化学反应,而非Fe3+,这主要是因为Fe3+常以不溶性的形态存在,导致其不易与NH2OH接触进行反应[73]。NO2-既可来源于硝化过程也可能来源于反硝化过程。在高土壤pH条件下施用铵态氮肥后易导致NO2-累积[74],而 NO2-可与Fe2+发生反应生成NO和N2O。此外,土壤NO2-也可与SOM发生亚硝基化反应 (图1E),且N2O产生量与SOM含量呈正相关[75]。
通常认为非生物学过程对N2O排放的贡献较少,如在21%和0%氧浓度下,粘壤土化学反硝化对N2O的贡献分别为6.33%和0.08%;同时,Ma等[42]发现添加NO2-后,灭菌土壤N2O排放占非灭菌土壤N2O排放比例少于1.1%。也有研究发现,添加NO2-后非生物因素对总N2O排放量的贡献较高,达31%~75%[80],这可能与土壤pH有关。
2 主要环境因子对N2O排放的影响
2.1 氮源
农田土壤施入的化肥氮约有70%以上是以铵态氮或酰胺态氮肥的形式进入土壤[76]。土壤氨氧化过程有中间产物NH2OH和NO2-的产生。这两种中间产物易于分解,因而在田间条件下的土壤中很难被监测到。特别是NH2OH,由于其极具活性,可经氧化作用产生NO2
-,发生化学作用产生N2O,也可与土壤有机质 (SOM) 发生反应。NO2-在土壤中也不稳定,同时由于土壤样品储运和处理过程较长等原因,也不易观察到其累积。但在实验室培养条件下,常常会发现NO2-的过渡性积累,且与N2O排放呈极显著正相关关系[77-80],这表明亚硝态氮作为产生N2O过程底物的重要性。
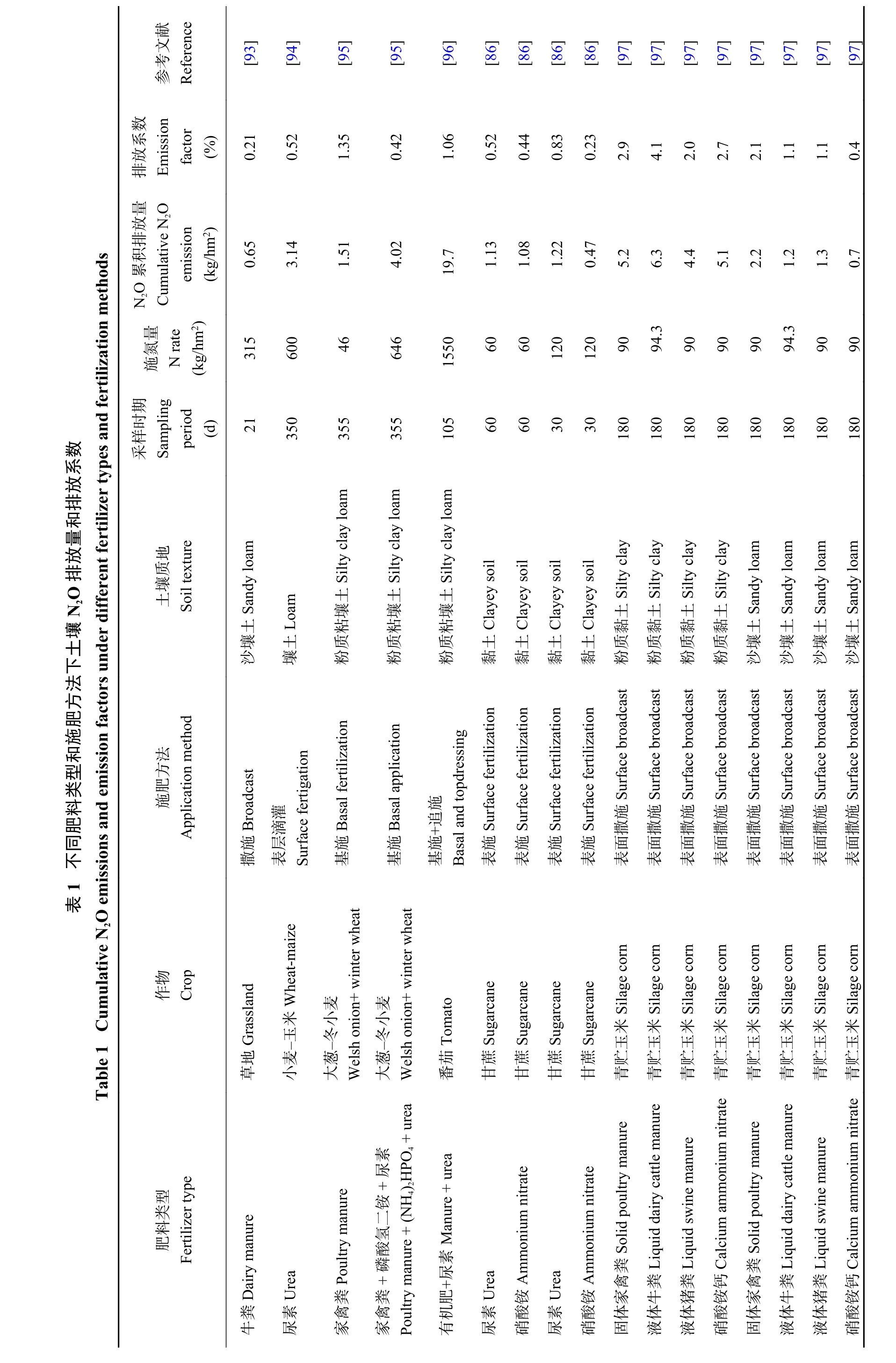
早期研究表明,施用化学氮肥能够增加农田土壤N2O的排放[81]。Shcherbak等[82]通过元分析发现,N2O排放随施氮量的增加呈指数而非线性增长。近年来研究发现,在覆膜水稻[83]和常规淹水水稻种植体系[84]土壤中,通过尿素氮深施可显著降低N2O排放。这可能由于深施后的尿素氮在土壤还原环境条件下多以铵态氮形式保存,且不易移动,降低了硝化和反硝化反应所需底物。对比不同肥料类型来看,通过田间试验大样本分析和模型显示,施用尿素产生的N2O排放量较硫酸铵高[85-86],这在室内培养试验中也得到类似结论[35]。同时,施用高效氮肥如包膜氮肥可显著降低土壤N2O排放[87]。Liu等[88]证实施用控释尿素土壤N2O排放系数显著低于尿素、硫酸铵和硝酸铵钙。Xia等[89]通过大样本分析发现施用控释氮肥可整体减少N2O排放达38.3%。与尿素[90]和硫酸铵[91]处理相比,土壤施用家禽粪肥可明显增加N2O排放量。大样本分析显示,与仅施用化学氮肥相比,有机肥增加N2O排放量平均高达32.7%[92]。有机肥与化肥混施 (表1) 或有机肥部分替代化肥[88]能够缓解土壤N2O排放。
?
许多研究显示,在我国北方石灰性土壤中硝化作用过程是产生N2O的主导途径[98-101],并推测这可能与该土壤低的有效性碳含量有关[101]。然而,在pH较高的情况下,化学氮肥施入土壤后由硝化作用导致的亚硝态氮过渡性积累,从而为硝化细菌的反硝化和反硝化过程进行提供底物,或许更为重要。在高铵态氮投入的中性土壤中,氨氧化细菌 (AOB)主导控制着硝化作用的活性[102],而氨氧化古菌(AOA) 通常在低NH3浓度的酸性土壤中发挥显著作用[103],这主要是由于AOA和AOB对底物 (NH3和O2) 的亲和性 (Km) 存在较大差异有关[104]。Hink等利用1-辛炔 (C8H14) 区分AOA和AOB对N2O产生量的相对贡献证实[105],添加铵态氮后AOB对土壤N2O总产生量起主导作用 (贡献比例为81%~86%),而未添加铵态氮的土壤中,AOA对N2O的贡献率为47%~58%,AOB对N2O的贡献可以忽略。因此,在施入或添加外源铵态氮造成NH3浓度较高时,AOB主导了硝化作用的进行,而当NH3主要来源于土壤的矿化作用时AOA是硝化作用过程进行的主要执行者。同时,长期向土壤中添加不同形态的氮素也会显著影响硝化微生物基因丰度。Zhou等[106]在连续进行44年施肥试验的草地土壤中发现,添加有机氮显著增加了AOA的amoA基因拷贝数,而AOB的amoA基因拷贝数在添加无机氮的处理中出现增加,且AOA/AOB比值随有机氮添加量的增加而增大。Ai等[107]在农田土壤中也发现,长期施用氮肥(尿素) 能显著增加土壤硝化潜势和AOB的种群丰度,而施用有机肥增加了AOA的种群丰度。这可能与AOA优先氧化有机氮 (来源于有机肥或土壤有机质) 矿化出来的氨有关,特别是在氨氧化速率超过矿化速率时,AOA对氨具有很高的亲和性[108]。而对于反硝化作用过程来说,增加土壤中NO3-通常会造成N2O/N2比值升高[109],这主要是由于NO3-作为末端电子受体 (terminal electron acceptor,TEA) 会被优先利用,且高NO3-含量也会抑制N2O还原酶活性[110]。
2.2 碳源
土壤碳源含量主要受耕作、有机肥投入、根系分泌物和作物残渣投入 (如秸秆) 等因素影响。Badagliacca等[111]研究发现,免耕措施下N2O排放显著高于传统耕作,这可能与免耕增加土壤有效性碳含量有关[112]。有机肥的施用显著增加土壤反硝化潜势。这主要是因为田间施用有机肥或其他有机物料后显著增加了土壤异养微生物的呼吸,同时有机物料中有效性碳为反硝化微生物提供了电子供体,促进了反硝化作用的发生。在植物根系根区内可通过根际沉积作用向根际土壤分泌一些低分子量碳的化合物,如糖类、有机酸和氨基酸等[113]。这些低分子量有机物极易分解,结合土壤根系的呼吸作用,造成根际土壤N2O产生量高于非根际土壤[114-115]。Giles等[116]对不同低分子量有机物研究发现,葡萄糖、谷氨酰胺和柠檬酸均显著增加N2的释放,且柠檬酸与葡萄糖和谷氨酰胺相比显著降低了N2O的排放。
目前,秸秆还田被认为是增加土壤肥力、提高土壤健康的农田管理措施之一。秸秆施入后主要通过影响土壤碳、氮有效性及土壤通气性进而间接影响N2O排放。Kravchenko等[117]研究发现,多数情况下秸秆施入可提高土壤N2O排放,并抵消土壤碳储存的气候效益。如在设施菜田土壤中,田间施用玉米秸秆能够显著增加N2O通量排放[118];在小麦-玉米轮作的粮田种植体系,小麦秸秆施用后可显著增加玉米季N2O排放量[119]。秸秆还田在不同农田土壤土地利用方式下对N2O排放的影响也不一致。Shan等发现秸秆还田在休耕地、旱地土壤中显著增加N2O排放,而在稻田中N2O排放显著降低[120]。与施用有机肥相比,通过秸秆深施可显著降低小麦季土壤N2O排放量[121]。Li等[122]通过15N示踪技术直接证明了作物秸秆施用后在不同水分条件下反硝化过程是土壤N2O排放的主导途径。然而,作物秸秆施用后并未发现土壤的反硝化微生物数量出现明显增加。Philippot等[123]研究发现,短期添加小麦秸秆对土壤反硝化菌基因丰度影响有限。同时,短期添加小麦、油菜和苜蓿秸秆也没有改变反硝化菌的数量,其反硝化作用与反硝化菌群落丰度呈解耦关系,与呼吸作用显著相关[124]。作物秸秆碳氮比是影响氮素释放的重要因子,可调控土壤氮素的固定和释放[125]。一般认为,高碳氮比植物残体施入土壤后可增加氮素固持进而有利于土壤肥力的提高,而低碳氮比植物残体会增加N2O的排放[120,126]。Chen等[14]利用大样本分析发现,作物秸秆碳氮比 < 45时对土壤N2O排放有显著正效应,而在碳氮比 > 100时略有负效应。同时,作物秸秆对土壤N2O排放的影响与土壤理化性质,特别是土壤水分和质地紧密相关。如,在低碳氮比土壤中施入动物有机肥可导致N2O排放量增加[127],而在高土壤碳氮比下,氮素气态损失主要以N2为主[128]。类似地,增加土壤有效性碳与硝态氮的比例可促进N2O还原为N2[129]。此外,大多数研究已证实,土壤施用生物炭能够显著减少N2O的排放[130-131],这主要与生物炭和土壤性质以及它们的交互效应有关。生物炭可通过改变土壤物理 (气体扩散、团聚体、保水性等)、化学 (pH、Eh、有效性有机和矿质态氮、溶解性有机碳等) 以及生物学性质(微生物种群结构、微生物量及活性、与氮循环相关酶活性等) 影响N2O的排放[132]。如生物炭通常具有高C/N比,并含有小分子量有机化合物,其多孔的结构适宜微生物生长并利于土壤氮素的固定,进而减少氮素有效性,降低土壤N2O的排放。同时,具有碱性特性的生物炭可用来改良酸性土壤,提高土壤pH。相关研究也已证实,生物炭可增加土壤微生物nosZ基因丰度,进而促进N2O的还原[133-134]。
2.3 水分含量
土壤水分是环境驱动力中最重要的因素之一[8]。水分影响土壤微生物细胞代谢活性和养分的运输[135],对硝化和反硝化作用过程具有决定性影响[136]。在土壤水分饱和区或淹水条件下,高土壤水分含量阻碍气体扩散,使土壤形成嫌气环境,增加土壤反硝化潜势和速率。同时,在该条件下,硝化作用进行得较为缓慢,参与反应的硝化微生物在植物根系周围的氧化区域具有较高活性[137]。在土壤水分非饱和区,其易于硝化作用的进行,该水分条件下反硝化作用主要发生在土壤团聚体和土壤微域环境中[138]。以土壤孔隙含水量 (WFPS) 作为判定N2O主要来源的指标已有大量研究。一般认为,硝化作用在30%~60% WFPS下是产生N2O的主要来源,而在土壤水分含量高于70%时N2O主要源于反硝化过程[138]。然而,在以硝化过程为主导的土壤环境中,N2O的排放总量远低于以反硝化过程主导的土壤环境[34]。土壤灌水后N2O含量迅速增加主要是反硝化作用的结果[139]。农田土壤玉米播种灌水后WFPS由20%迅速增加至80%,其产生的N2O剧烈排放可能主要来源于反硝化作用过程[140]。设施菜田土壤N2O排放主要集中在基施有机肥并灌水后的1周内,N2O排放也主要受反硝化作用影响[119]。此外,施用有机肥并灌水显著增加了分子态氮 (N2) 的损失[141]。
水分含量影响土壤硝化和反硝化微生物的活性。Uchida等[142]发现在土壤淹水后的几小时内(3 h),反硝化基因 (nirK,nirS,nosZ) 在mRNA水平0—1 cm土层土壤中显著增加并达到峰值,也就是说当水分含量造成土壤氧气浓度发生改变时,反硝化细菌会迅速做出反应以适应新的环境条件,表明降雨或灌溉后导致N2O排放峰的出现与反硝化过程有关。与此类似的是,Riya等[143]研究表明,在土壤淹水消失前48 h添加NH4+后,amoA的mRNA拷贝数在5—10 mm土层土壤中虽有增加,但不显著,而nirK的mRNA转录水平在淹水消失6 h后急剧增加。然而,与上述观点不同的是,Theodorakopoulos等[144]发现,随着土壤WFPS上升,并未发现在0—10 cm土层土壤中反硝化基因转录数 (mRNA) 增加,而显著增加了细菌amoA的转录数,且其与N2O排放呈极显著正相关,这可能与土壤中低氧气含量和铵态氮有效性增加有关。
2.4 氧气
土壤氧气含量是驱动生物地球化学循环的关键因素之一,也是影响硝化作用和反硝化作用对N2O排放相对贡献的决定因子之一[145]。在自然生态系统中,空气对流和气体扩散影响土壤中的氧气含量;同时,异养微生物呼吸作用或微生物耗氧分解以及硝化作用的发生也会减少土壤氧气浓度[146]。Groffman等[147]认为土壤“热点”(hot spots) 的形成主要是因为水分含量的提高和有机肥施用后增加了土壤有效性碳氮的富集,进而影响了氧气浓度在土壤中分布。在氧气受限而有效性碳充裕的土壤环境下,其“热点”区域N2O的排放以反硝化作用为主导途径。氧气浓度在土壤中的非均质性分布,也是导致N2O产生或排放出现差异性的重要原因[148]。
氧气浓度影响硝化、反硝化作用对N2O排放的贡献。Khakil等研究发现,在低氧浓度下 (0.35 kPa)反硝化作用是产生N2O排放的主导途径。随土壤厌氧环境的形成,硝化作用对N2O排放的贡献明显下降[148]。Zhu等[34]研究发现,在低氧条件下氨氧化和硝化细菌的反硝化作用是产生N2O的主要来源。其中,在0.5%和3%(体积比) 氧浓度下,硝化细菌的反硝化作用对N2O排放的贡献为34%~66%。这可能主要是由于在低氧条件下,氨氧化细菌 (AOB) 的群落组成发生改变,一些AOB物种能够进行反硝化作用,如亚硝化单胞菌和类亚硝化单胞菌在低氧条件下丰富度增加,同时异养反硝化作用对N2O的贡献量主要受氧气和有效性碳含量的影响[149]。需要注意的是,部分硝化微生物在利用亚硝酸盐而不是氧气作为电子受体时,氧气仍是氨氧化过程反应发生所不可缺少的条件[137]。
2.5 土壤 pH
土壤pH可直接通过影响生物[150]和非生物学过程调控N2O的排放。pH也能够改变土壤底物有效性,如低pH会降低土壤矿质氮和有机碳的可利用性进而间接影响N2O排放[151]。大样本分析证实,降低土壤pH会增加N2O排放[152]。Sun等[153]发现,在草原和森林土壤中N2O/N2比值随土壤pH下降而升高。在我国农田土壤中,由于过量施用氮肥,土壤pH明显下降[154],或将大幅度增加N2O排放。
硝化作用过程对土壤pH的响应可能较为敏感,这主要与pH影响NH3有效性 (AMO的底物) 有关[;pKa= 9.25,25℃][155]。每改变1个pH单位NH3浓度的变化将近10倍[156]。硝化速率通常随土壤pH的增加而上升[157-158],其培养的硝化细菌最适pH范围为7.5~8.0[157]。对于氨氧化微生物而言,增加土壤pH可以增加氨氧化古菌 (AOA)amoA基因丰度,而对氨氧化细菌 (AOB) amoA基因丰度没有影响,甚至有些减少[159-160],但也有人认为土壤pH增加可以提高AOB的丰度,而对AOA没有影响[161-162]。大量研究发现酸性土壤硝化作用的主要贡献者为AOA[163-165],这与AOA在酸性条件下对NH3具有较高的亲和力有关[166]。研究表明,pH与硝化细菌的反硝化过程产生的N2O呈正相关关系,而与反硝化作用产生的N2O呈负相关[167]。Cheng等[168]研究发现,好氧条件下,当土壤pH约低于4.4时反硝化作用主导N2O的产生,降低土壤pH能够显著增加反硝化作用对N2O的贡献,这可能与真菌反硝化有关[169-171]。曹文超等[129]在菜田土壤中研究发现,增加土壤pH会提高土壤反硝化速率,并促进N2O还原为N2,这与Richardson等[81]发现的结果相一致,即在碱性环境下,反硝化主要产物为N2,N2O产生量较少。原因主要是低pH会干扰反硝化过程中N2O还原酶组装 (nosZ基因的表达),降低酶的活性[27,172],导致反硝化作用N2O排放量增加和N2O/(N2O + N2) 比值增大。此外,土壤pH也会显著影响非生物学过程对N2O的贡献[66]。如羟胺 (NH2OH) 在pH低于5.95条件下以较游离NH2OH更稳定的质子化形态 (NH3OH+) 存在,导致其有效性降低,进而减少了N2O排放。
2.6 温度
在全球气候变暖的大背景下,温度对N2O排放的影响十分重要。在高纬度地区,土壤温度是影响N2O排放的主要因子。如在我国三江平原地区不同施氮处理条件下土壤温度可解释N2O排放16%~33%的变异[173]。温度可直接和间接影响土壤N2O排放。直接调控主要表现在对土壤微生物生长和酶活性的影响,如降低土壤温度能够抑制土壤生物活性,进而减少N2O产生[174]。其间接调控主要是通过微生物活动改变土壤氧气和有效性碳的含量[175-176]。此外,氧气在土壤溶液中的溶解和释放也受温度的间接影响[177]。
温度影响土壤微生物反应活性,进而影响N2O排放。对于氨氧化微生物而言,与20℃相比,AOB和AOA的生长在30℃和40℃下均受到抑制[59]。Maag和Vinther[174]发现提高土壤温度可降低硝化作用对N2O的贡献,而增加反硝化过程N2O的产生量。Davidson等[177]也发现,随土壤温度提高,反硝化作用产生的N2O排放量增加。这可能是因为升温后增加了微生物的呼吸作用,造成土壤微域氧气亏缺,为反硝化微生物活动创造了厌氧条件[178],同时升温也增加了土壤反硝化活性[179]。许多研究表明,当土壤温度在10~35℃时,每升高10℃,土壤反硝化的活性可提高1.5~3.0倍[180-181]。类似地,Saleh-Lakha等利用Pseudomonas mandelii进行液体培养发现,其在10~30℃范围内反硝化作用活性随温度的上升而增加[182]。对于施用有机肥后的碱性土壤而言,提高土壤温度显著增加N2O排放,同时,与仅施用化学氮肥处理相比,有机肥施入增加了反硝化作用对N2O总排放量的贡献。这可能与提高温度后显著增加了反硝化nirS基因丰度有关[183]。然而,在酸性土壤中研究发现,30℃时施用有机肥处理的nirK基因丰度显著增加,而nirS在该温度下迅速降低[59]。
2.7 其他因素
土壤N2O排放随大气CO2浓度的增加而增加[184-185]。这可能与增加土壤反硝化作用对N2O的贡献有关。一方面,大气CO2浓度升高后减少了植物地上部 (如叶片) 的蒸腾作用,进而使土壤含水量增加;另一方面,高CO2浓度促进了植物地下部有效性碳的分配,如增加根系分泌物的释放等。二者均有益于反硝化作用的进行[184]。但也有研究认为N2O排放增加是由于硝化作用贡献增加的结果[186]。此外,CO2浓度提高后,真菌的活动使草地土壤N2O排放潜势增加了49%,同时,CO2浓度升高显著增加真菌基因丰度和真菌与细菌拷贝数比例[187]。
土壤结构也是影响N2O排放的重要因素之一,其主要影响土壤通气性和气体扩散,进而影响N2O的产生[188-189]。在农田土壤中,机械化操作导致土壤压实可造成土壤N2O排放量增加[190]。土壤动物活动也会影响N2O排放[191]。如大样本分析表明土壤大型动物蚯蚓活动增加N2O排放量达42%[192]。Zhu等研究发现蚯蚓和弹尾虫二者显著增加土壤N2O排放量[193]。除蚯蚓外,在许多生态系统中,蚂蚁生活所形成的蚁丘土壤是CO2排放的“热点”区域[194-195]。近年已有研究表明,蚁丘土壤N2O产生量显著高于相邻自然土壤,这主要是由于蚂蚁活动增加了土壤碳氮和粘粒的含量[196]。此外,不同蚁群种类形成的蚁丘土壤对N2O和CO2排放也有差异[177]。
3 农田土壤水肥管理对N2O排放的综合影响
农田土壤N2O的产生和消耗本质上是土壤碳、氮、氧等因子综合作用的结果,而田间水、碳、氮的管理措施是影响这些环境因子变化的主要因素 (图2)。综合田间土壤N2O通量监测结果,N2O峰值排放阶段是其排放的关键,该时期N2O累积排放量约占全年农田排放量的50%左右[197-198]。
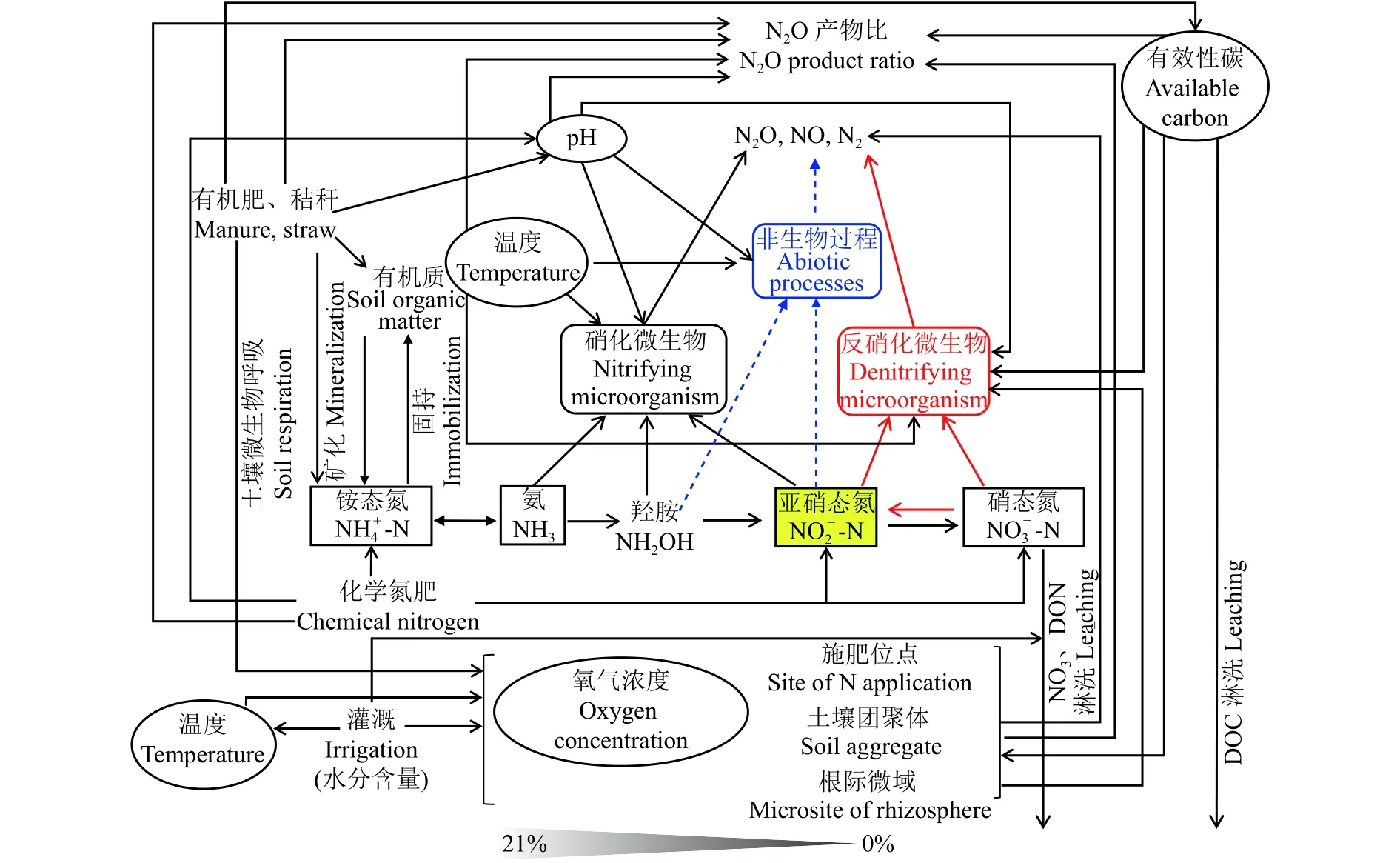
图2 农田土壤主要调控因子对N2O排放综合影响的概念图Fig. 2 Conceptual diagram of comprehensive influences of main regulatory factors on N2O emissions in soils
在农田土壤水肥管理中,有机肥施入和灌水是造成N2O剧烈排放的主要诱因。以设施菜田土壤种植体系为例,有机肥极大地促进土壤中异养微生物的呼吸作用,显著降低土壤孔隙中氧气浓度;同时大量灌水降低了土壤相对气体扩散系数 (RD),造成土壤出现低氧甚至厌氧的土体环境,尤其在土壤施肥位点、土壤团聚体及根际微域等区域中更易形成,这为反硝化微生物活动提供了厌氧条件。此外,土壤有机氮或有机肥矿化形成NH4+后,硝化作用对土壤氧气的消耗也有贡献 (NH3+ 2O2→NO3-+H2O + H+)。有机肥施入土壤后为反硝化微生物提供了其所需的有效性碳,保障了反硝化作用过程发生所需的能源。值得注意的是,与大田土壤相比,设施菜田土壤硝态氮含量已不是反硝化过程发生的限制因子,因此,在其施用有机肥导致土壤有效性碳含量增加的短期时间内,将极大地促进以反硝化作用过程为主导途径的N2O峰值排放。
在土壤含水量较低 (质量含水量25%左右) 条件下,铵态氮肥施入可增加硝化作用过程N2O的排放,但其整体排放系数较低 (1%左右),且长期施用铵态氮肥会造成土壤酸化。土壤pH值的改变又将影响土壤微生物种群结构的变化。以氨氧化微生物细菌 (AOB) 和古菌 (AOA) 为例,由于它们在不同的土壤环境条件下占据不同的生态位[199-201],其对土壤pH值的响应也不相同。一般认为,AOB在中性、碱性和高氮素投入的条件下是驱动硝化过程进行的主体,而AOA在酸性的自然生态系统中更能发挥作用[202]。同时,对于反硝化微生物来说,低pH阻碍N2O还原酶的形成[203],导致N2O产物比值增加。值得注意的是,非生物学过程在低pH土壤中对N2O的贡献不可忽略,特别是在过量施肥导致土壤出现NO2-累积时,非生物学过程对N2O排放的贡献更加明显。此外,温度也是影响农田土壤N2O排放的重要因子,在全球气候变暖的大背景下会受到更多关注。在农田土壤水肥管理措施中,硝化抑制剂、缓控释肥等高效施氮 (见2.1部分) 和间歇灌溉等技术的应用,有利于农田土壤N2O的减排。
4 结论与展望
综上所述,虽然土壤N2O产生途径有反硝化、硝化、硝化细菌的反硝化和硝化-反硝化耦合、真菌反硝化、化学反硝化,以及硝酸异化还原为铵等,但就一般农田土壤而言,生物反硝化过程或许仍然是N2O产生的主要途径。支持这一观点的依据既表现在环境因子和管理措施对农田土壤N2O排放的影响机理方面,而且也有分子生物学的证据。而硝化细菌的反硝化、硝化-反硝化耦合过程等,实质上也是生物反硝化过程的一部分。硝化过程本身对农田土壤N2O排放的直接贡献较小,在许多情况下可能仅仅是背景排放。然而,由于硝化过程为生物反硝化和化学反硝化提供了底物,特别是中间产物NO2-的释放,其对N2O排放具有很大的间接贡献。此外,真菌反硝化和化学反硝化在酸性土壤以及DNRA过程在高有机质和厌氧的土壤环境中,也对土壤N2O排放具有不可忽视的重要作用。合理管理有机肥和水分,辅之以硝化抑制剂的利用[204-205],可在很大程度上减少农田土壤N2O排放。
目前人们对农田土壤N2O的产生和消耗方面的认识还面临诸多挑战。主要表现在:首先,农田土壤N2O排放主要来自于自然降水后,特别是施肥后立即灌水而产生的排放峰。在这一阶段的N2O产生和消耗过程进行深入研究,对农田土壤N2O减排措施的提出具有重要意义。其次,降低土壤反硝化过程N2O/N2的产物比,是N2O减排的有效措施之一。为此,需要深入了解环境因子和调控措施对反硝化产物比的影响及其机理,以便为通过降低产物比实现N2O减排提供依据。再者,加强对N2O还原过程及其影响因素的了解,也具有重要意义。最后,在寻找与N2O排放有关的关键微生物方面人们遇到了很大挑战,这与反硝化微生物功能类群的广谱性和生理类型的多样性有密切关系,相信随着分子生物学和基因芯片技术的发展,这一问题能够逐步得到解决。与此同时,综合利用新技术方法,如平面光极技术[148]、Robot自动培养技术[206]、氦环境培养—气体同步直接测定技术[207]、宏基因组测序技术等进一步探究土壤物理、化学和生物学因素对氮素转化过程、N2O关键位点排放及关联微生物多样性的影响,构建氮素平衡和相关N2O排放模型,可进一步为N2O排放机制及相关影响因素提供理论依据。此外,通过对反硝化过程中N2O/N2产物比产生规律及其影响因素的深入了解,将为利用田间N2O监测数据定量估算氮的反硝化损失提供一种新的途径。
猜你喜欢
湘潮(上半月)(2021年10期)2021-12-02 02:09:38
昆明医科大学学报(2021年5期)2021-07-22 07:32:48
公民与法治(2020年15期)2020-09-25 02:57:54
知识经济·中国直销(2018年1期)2018-01-31 01:52:37
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
商周刊(2017年6期)2017-08-22 03:42:37
中国环境科学(2016年3期)2016-02-08 15:07:14
应用海洋学学报(2014年2期)2014-11-26 01:20:46
中国酿造(2014年9期)2014-03-11 20:21:15
生物加工过程(2013年1期)2013-03-11 18:31:35
