
川西平原灌区不同水旱轮作模式周年土壤呼吸特征
2019-11-07 06:43杨志平任万军
生态学报 2019年18期
傅 勇, 王 淘, 杨志平, 周 伟, 刘 琦, 任万军, 陈 勇
四川农业大学农学院/作物生理生态及栽培四川省重点实验室, 温江 611130
工业革命以来,由于人类活动的影响,大气CO2浓度急剧升高,增加了约 32%,全球正经历着变暖的危机[1,2]。农田生态系统是陆地生态系统的重要组成部分,占全球陆地面积的10.5%,是全球碳循环中最活跃的部分,CO2占人为温室气体排放量的21%—25%,且受人类干扰频繁[3]。土壤碳库是大气碳库的2倍,土壤中的CO2浓度发生微量的变化,就会对大气中CO2浓度造成巨大影响[4, 5]。土壤呼吸是农田生态系统土壤碳排放的主要途径,土壤通过呼吸作用向大气释放CO2的年通量为5.0×1016—7.5×1016g,是土壤碳素以CO2形式回归大气碳库的主要路径,占陆地生态系统与大气之间碳交换总量的2/3,对气候变化和碳循环起着不可忽视的重要作用[6, 7]。土壤呼吸作为碳循环的核心问题已引起了国内外学者的密切关注和广泛研究,并取得了大量的研究成果。
近年来,我国农田生态系统土壤呼吸的研究主要集中在不同作物类型、土地利用方式或栽培措施下土壤呼吸的时空变异特征及调控因素的研究,研究对象涉及小麦、玉米、水稻等主要农作物[8- 10]。很多研究表明,农田土壤呼吸受环境因子共同调控,尤其对土壤温度和土壤含水量的响应十分敏感[11-12]。也有研究表明,与土壤温度相比,土壤水分对土壤呼吸季节变化的影响更为重要[13]。但前人关于作物系统的研究对象多为旱地轮作系统或单一作物系统,缺乏对稻田水旱轮作系统周年土壤呼吸的探索。水旱轮作是我国南方稻区十分重要的种植制度,但由于旱、水土壤环境转换,土壤理化性质、土壤有机质矿化分解速率、微生物种类、微生物数量和活性、根际生物量等发生剧烈变化,土壤呼吸随之受到极大的影响[14,15]。为了从周年种植模式角度探讨不同水旱轮作系统周年土壤呼吸特征及其影响因素,本研究基于蒜-稻、麦-稻以及油-稻三种川西平原灌区重要的水旱轮作模式的定位试验,探明不同轮作模式及其不同生长季土壤呼吸的变化趋势和差异特征;探明土壤呼吸与土壤温度、土壤含水量等生态因子的关系,以期为水旱轮作系统减排降耗提供理论依据。
1 材料与方法
1.1 研究区域概况
试验设在四川省崇州市四川农业大学现代农业研发基地(30°33′N,103°38′E),位于川西平原都江堰灌区,属亚热带湿润季风气候区。该区年平均气温15.9℃,平均降雨量1012.4 mm,平均日照时数为1161.5 h。试验地基础养分含量为土壤有机质31.58 g/kg,全氮1.85 g/kg,速效磷11.81 mg/kg,速效钾85.48 mg/kg。
1.2 试验设计与管理
试验地采用水旱轮作方式进行试验,已连续进行3年的水旱轮作模式研究。采用单因素随机区组设计,设置3个水旱轮作模式处理,分别为小麦-水稻(WR)、大蒜-水稻(GR)和油菜-水稻(RR),每年的处理保持一致, 3次重复,共9个小区,小区面积为26 m2(4 m×6.5 m)。除水稻季施肥采用氮肥后移技术外,其余各作物施肥及管理方案均按照农民常规方法进行。本研究的时间范围为2017年5月—2018年5月,为1个完整的水旱轮作周期,其中2017年5月—9月为水稻季,2017年9月—2018年5月为旱作季,轮作周期的各作物播栽期及播栽密度见表1,耕作、施肥及管理措施见表2。

表1 各作物播栽期及播栽密度
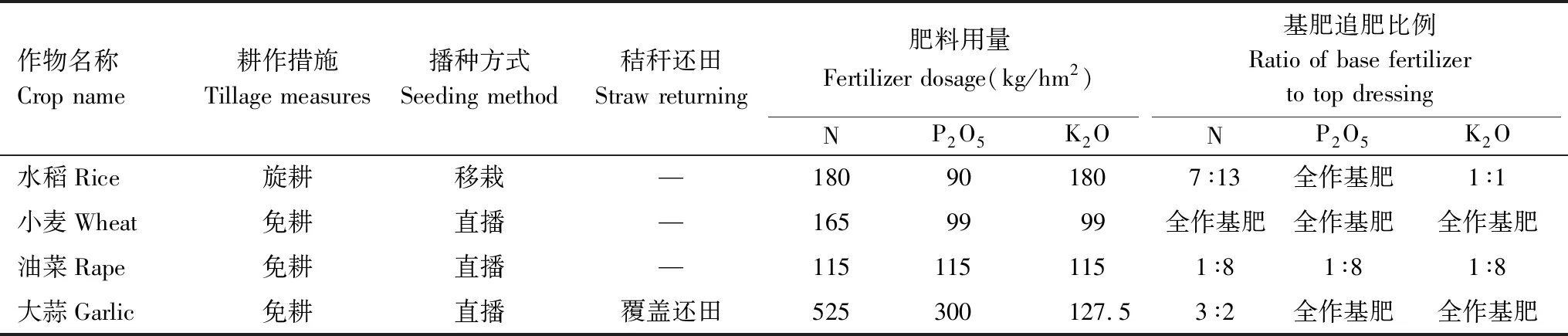
表2 不同作物耕作、施肥及管理措施
1.3 测定指标与方法
1.3.1土壤呼吸测定
土壤呼吸测定于2017年5月25日开始,每隔30 d选择晴天(遇雨适当延后)进行土壤呼吸测定[16,17]。其中,土壤呼吸季节动态选择取样当天9:00—11:00进行取样,土壤呼吸日动态取样从9:00至18:00每隔3 h取样一次;18:00至次日6:00每隔4 h取样一次。水稻移栽后,于各小区行间分别放置1个由PVC材料制成的土壤呼吸气体收集装置,该装置参照王维钰等[18]的设计,并根据田间取样环境进行了一定的改进,形成腔室高25 cm,直径为16 cm,插入土壤5 cm。腔室顶部为开放结构,开放部分直径为11.5 cm,确保腔室内外部环境条件一致。取样时加盖密封,盖顶开有两个小孔,分别用于安装温度计和连接橡胶软管(橡胶软管连接至PVC腔室中心)。取样时加盖密封后,盖顶用橡胶软管连接注射器,采样时间为加盖后0、10、20和30 min,取60 mL气体置于真空袋中待测,取样后立即用铁夹夹住橡胶软管保持密闭。样品浓度用岛津2010Plus气相色谱仪检测分析,通过对4个气样浓度进行线性回归,计算气体排放速率。气体排放速率计算公式如下[19]
式中,F为CO2排放速率,mg m-2h-1;ρ为标准大气压下的CO2密度,为 1.97 kg/m3;T为采样过程中腔室内的平均温度,℃;H是腔室的净高度,m;dC/dt是采样气体浓度的变化率。
土壤呼吸累积排放量计算公式如下:
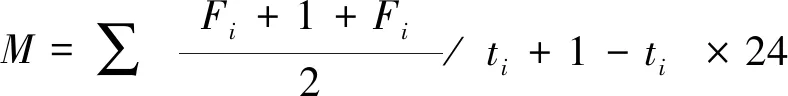
式中,M为土壤CO2排放总量(kg/hm2);F为CO2排放速率(mg m-2h-1);i为第i次测定;(ti+1-ti)为相邻两次测定间隔天数。
1.3.2水热生态因子测定
于每月土壤呼吸取样时,同步测定土壤温度、土壤含水量,稻季测定水层高度。土壤温度采用便携式温度测定仪测定,分0—10 cm和10—20 cm测定。各小区按五点取样法采集0—20 cm土样,采用烘干法测定土壤含水量;田间水层高度用直尺测量。
1.3.3土壤呼吸与水热因子拟合
土壤呼吸速率和土壤温度、土壤含水量和水层高度分别采用指数函数模型和一元二次模型拟合[20,21]:
Y=aebT
Y=X2+X+c
温度敏感性系数Q10代表土壤呼吸的温度敏感性,计算公式如下[17-18]:
Q10=e10b
式中,Y为土壤呼吸速率(mg m-2h-1),T为平均温度(℃),a为0℃时土壤呼吸速率,b为温度反应系数,X为土壤含水量(%),c为截距,Q10指温度升高10℃时土壤呼吸速率变化的倍数。
1.3.4数据处理
采用SPSS13.0进行数据处理,并用Microsoft Excel 2010作图。
2 结果分析
2.1 不同水旱轮作模式土壤呼吸速率季节变化特征
图1表明,三种模式土壤呼吸速率季节变化规律趋于一致,周年呈现双峰型变化趋势,6—9月土壤呼吸速率显著高于其余月份。三种模式土壤呼吸速率分别在6月和9月达到峰值,其中GR和RR模式在6月达到全年呼吸速率最大值,分别为63.1 mg m-2h-1和61.2 mg m-2h-1,WR模式则在9月达到全年呼吸速率最大值,为36.82 mg m-2h-1;而三种模式土壤呼吸速率最低值出现在1月,GR、WR、RR模式土壤呼吸速率最低值分别为3.89 mg m-2h-1、4.49 mg m-2h-1和1.42 mg m-2h-1。不同模式之间,GR模式全年平均土壤呼吸速率显著高于其余两种模式,而以WR模式最低。
2.2 不同作物生长季节土壤呼吸累积排放量
图2表明, GR、RR和WR三种模式周年土壤呼吸累积排放量分别为193.36 kg/hm2、157.28 kg/hm2和160.27 kg/hm2,其中累积排放最高的GR模式分别比RR和WR模式高17.11%和18.5%。季间空闲期,WR模式土壤呼吸累积排放量为41.84 kg/hm2,显著高于其余两种模式;从不同作物生长季来看,GR、WR、RR三种模式均在稻季累积排放量最高,分别占周年累积排放量的49.28%、48.11%和58.33%; GR、WR、RR三种模式稻季土壤呼吸累积排放量较旱季分别高16.43 kg/hm2、35.88 kg/hm2、53.68 kg/hm2。在稻季,GR模式和RR模式的土壤呼吸累计排放量显著高于WR模式,两种模式分别比WR模式高出25.93%和23.56%;而旱季GR模式土壤呼吸累计排放量达78.5 kg/hm2,显著高于WR模式和RR模式,分别比WR模式和RR模式分别高出98.23%和98.08%。

图1 不同水旱轮作模式周年土壤呼吸速率特征 Fig.1 Characteristics of soil respiration rate in different paddy-upland rotation
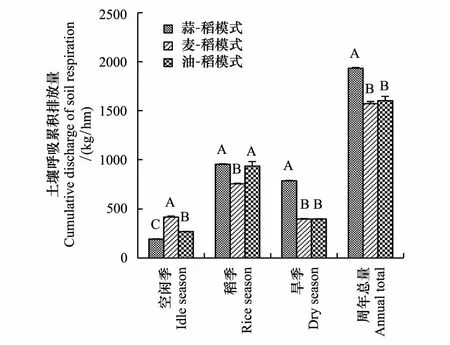
图2 不同水旱轮作模式土壤呼吸累积排放量 Fig.2 Cumulative emission of soil respiration in different paddy-upland crop rotation system同组中不同英文字母分别代表多重比较下在P<0.01水平下差异达显著水平
2.3 不同水旱轮作模式土壤呼吸速率日动态变化特征
如图3所示,三种模式土壤呼吸速率昼夜变化主要呈现单峰型变化,总体表现为白天高,夜晚低,具有一定波动性。各模式土壤呼吸速率在12:00—15:00达到最大值,其中WR模式在2月、10月和12月最大值出现的时间滞后于其余两种模式;三种模式土壤呼吸速率在2:00—6:00出现最低值,不同季节各模式最低值出现的时间略有差异。GR、RR模式土壤呼吸速率日变化幅度在6月出现最大值,分别为31.11 mg m-2h-1和52.52 mg m-2h-1,WR模式土壤呼吸速率日变化幅度在8月出现最大值,为28.29 mg m-2h-1;GR和RR模式在2月出现日变化幅度最小值,分别为6.52 mg m-2h-1和3.56 mg m-2h-1,而WR模式日变化幅度最小值出现在12月,为7.63 mg m-2h-1。GR模式各月份土壤呼吸速率日动态波动较小,而WR模式和RR模式土壤呼吸速率日动态变化相对剧烈;WR模式土壤呼吸速率在2月和4月的15:00—18:00之间分别下降54.34%和58.07%;RR模式土壤呼吸速率在4月和6月的15:00—18:00之间分别下降65.21%和30.22%。

图3 不同水旱轮作模式土壤呼吸速率日动态特征Fig.3 Daily dynamic characteristics of soil respiration rate in different paddy-upland crop rotation system
2.4 土壤呼吸速率与土壤含水量、水层高度及土壤温度的关系
通过对土壤呼吸速率与旱季土壤含水量、稻季水层高度进行回归分析,结果表明,在旱季,土壤呼吸速率与土壤含水量没有显著相关性(P>0.05),但稻季受水层高度影响较大,其相关性达到显著水平(P<0.05)。通过对土壤呼吸速率与土壤温度的回归分析发现,不同模式土壤呼吸速率与0—10 cm、10—20 cm土壤温度均存在极显著的相关性(P<0.01),即土壤呼吸速率随着土壤温度的升高而增高。不同模式土壤呼吸速率对土壤温度的敏感性有所差异,表现为GR>WR>RR;从不同土层来看,三种模式Q10均表现为0—10 cm>10—20 cm,表明随土层的深入,土壤呼吸速率与土壤温度的敏感性逐渐减小。不同模式不同土层深度Q10变化范围为1.15—2.72,以GR模式0—10 cm最高,RR模式10—20 cm最低。
3 讨论
3.1 不同水旱轮作模式对土壤呼吸速率影响
本研究结果表明,三种模式土壤周年呼吸累积排放量表现为GR>RR>WR。研究表明[22,23],秸秆还田能够明显提高农田土壤碳固定,有效提高土壤有机碳及活性有机碳组分含量,而土壤有机质是微生物进行分解活动排放CO2的物质基础[24]。前人研究表明[25],由于“启动效应”的存在,土壤中碳的投入会导致土壤矿化的短期增加,从而导致碳流失。在本研究中,GR模式由于旱季进行秸秆覆盖还田,增加了外源有机质的输入,提高了土壤有机碳含量,为微生物活动提供了大量底物,同时加速土壤矿化,增加了土壤呼吸速率,从而使GR模式土壤呼吸周年累积排放量显著高于WR、RR模式。此外,王春新、张庆忠[26, 27]等研究表明,施氮量对土壤呼吸具有显著影响,在本研究中,GR模式旱季施氮量远高于WR、RR模式,GR模式旱季较高的氮投入不仅促进了大蒜地上和地下部分生物量的增加,同时残留的氮素在稻季得到利用,增加了GR模式稻季生物量的累积,从而间接提高了GR模式土壤呼吸速率,这也可能是导致GR模式土壤呼吸累积排放量高于其余两种模式的原因之一。
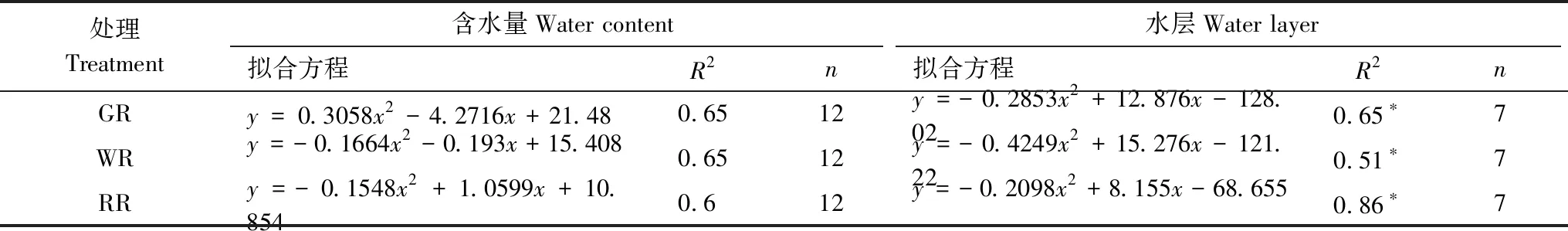
表3 土壤呼吸速率与含水量、水层的关系
*—P<0.05 ;**—P<0.01
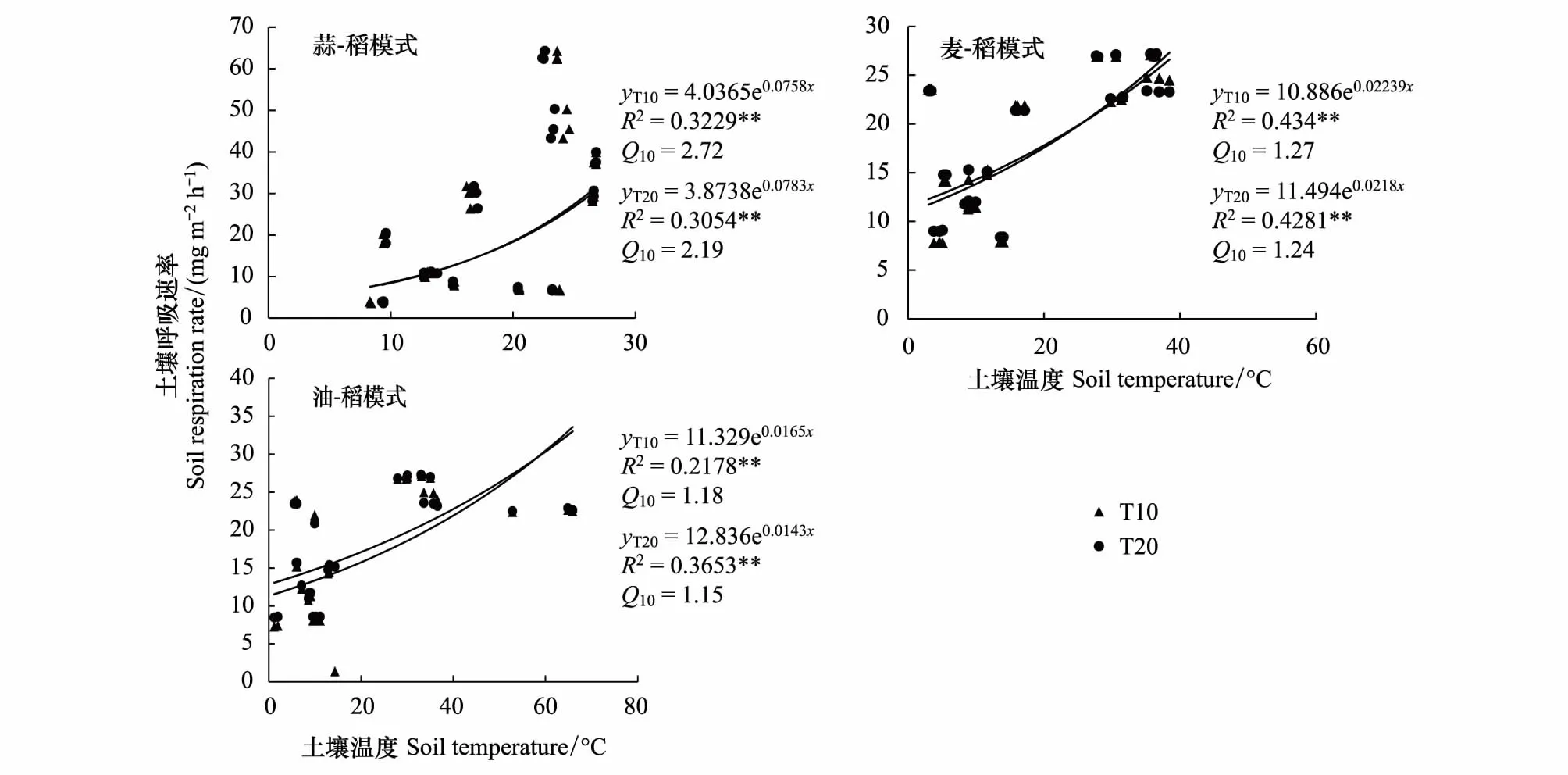
图4 土壤呼吸速率与土壤温度的关系Fig.4 Relationship between soil respiration rate and soil temperature
从土壤呼吸速率季节变化特征来看,水稻生长季土壤呼吸速率要显著高于其余季节。在水稻生长季中,6月底处于水稻分蘖末期晒田阶段,植株根系生长旺盛,根际微生物活跃,土壤呼吸速率增强;此外由于没有了水层的阻隔,土壤中闭蓄态CO2得以释放[28, 29],也使得土壤呼吸速率增大。9月水稻收获后,田间含水量降低且不受植株遮阴影响,土壤呼吸速率又出现一个峰值。GR模式大蒜播种后,受秸秆覆盖还田和氮肥投入影响,GR模式土壤呼吸速率高于其余两种模式。旱季作物生长过程中,随着气温的逐渐降低,三种模式土壤呼吸速率逐渐下降,至1月降至最低,之后土壤呼吸速率缓慢回升。但三种模式土壤呼吸速率在12月份有所上升,这可能是由于作物生育进程和地下生物量的增加从而增加了土壤微生物的活性,进而提高了土壤呼吸速率。
三种水旱轮作模式土壤呼吸日动态主要表现为单峰形式,土壤呼吸速率总体白天高于夜间,峰值出现在12:00—15:00之间,最低值则出现在凌晨2:00—6:00之间,这与高会议、韩广轩等[30,31]研究结果一致。但4月WR模式22:00土壤呼吸速率高于18:00土壤呼吸速率,2月GR模式呼吸土壤速率最低值出现在22:00,与气温和土壤温度变化规律不一致,这与陈亮等[32]在黄河三角洲滨海湿地的研究结果类似,这可能与温度降低后地表受霜降形成的水雾有关。GR模式土壤呼吸速率日均值在8月、10月和12月高于其余两种模式,这主要是由于秸秆覆盖还田增加了土壤的有机质含量,为微生物活动提供了大量底物,加速了土壤矿化,从而提高了土壤呼吸速率。此外,各模式不同季节土壤呼吸速率日动态的平均值接近9:00的土壤呼吸速率,与季节动态的取样时间相吻合,这与前人研究结果相一致[33]。
3.2 土壤呼吸速率与温度、土壤含水量及水层高度的关系
土壤温度是影响土壤呼吸速率的主要因素之一,研究表明[8,34],土壤温度在一定范围内增加可以提高土壤微生物活性,从而影响土壤呼吸,但当土壤温度超过一定范围,抑制了土壤微生物活性,使土壤呼吸速率降低。本研究结果表明,土壤呼吸速率与土壤温度呈指数正相关关系,这与崔海[35]、张俊丽[36]等研究结果一致。研究者们通常用Q10来表示土壤呼吸速率与土壤温度之间的关系,前人研究结果表明[37],Q10值变化范围一般在1.3—3.3之间。在本研究中,三种模式Q10值变化范围在1.15—2.72之间,与Raich等[37]的研究结果类似。不同模式间,GR模式Q10值最高,表明GR模式土壤呼吸速率与土壤温度敏感性最高,这可能是与GR模式采用秸秆覆盖还田有关。
土壤含水量是除土壤温度外,影响土壤呼吸速率的又一关键因素。土壤含水量通过调节土壤通气状况,影响微生物活性以及调节土壤氧化还原状况来调节土壤呼吸;土壤水分变化对温度的影响(通过水分有效性驱动的感热通量和潜热通量的变化)也间接影响土壤呼吸。在本研究旱季作物生长过程中,土壤呼吸速率与土壤含水量没有显著相关性, GR、WR、RR模式分别在含水量达到22.57%、17.98%、19.44%时,各模式土壤呼吸速率达到最大值。土壤含水量对土壤呼吸的影响十分复杂,往往同时受与土壤温度的协调情况影响,目前关于土壤含水量与土壤呼吸速率关系的研究尚没有定论。多数研究者认为,土壤含水量在一定范围时,随着土壤含水量的增加,土壤呼吸速率相应上升,当土壤含水量超出这个范围时,则会抑制土壤呼吸速率。王雪等[38]研究表明,土壤水分与土壤呼吸速率呈倒“U”型相关关系,即土壤水分含量较低时,土壤水分的增加可以促进土壤呼吸作用,但土壤含水量达到一定程度后,土壤水分的增加则抑制土壤呼吸作用,这可能是由于土壤在干旱阶段,微生物活动由于受到水分的限制;而在土壤处于湿润阶段时,有机物的分解过程从好氧转变为厌氧,而厌氧分解速率仅为好氧分解速率的30%—40%[39]。丁新宇等[40]研究指出,在不同土壤条件下,土壤含水量呈现完全相反的相关性,在干旱阶段,土壤呼吸速率与土壤含水量正相关,但在湿润阶段土壤呼吸速率与土壤含水量则呈负相关。
在水稻生长季,淹水状态下土壤呼吸速率受水层深度制约[41]。本研究结果表明,土壤呼吸速率与稻田水层高度呈显著负相关,这与武文明等[28]在西双版纳地区的研究结果一致。 由于水层的存在,土壤呼吸受到阻隔,同时水层能够溶解部分CO2;此外,水层深度的变化还会影响土壤温度、有机质分解、微生物活性和土壤动物的数量等其他因子,进而影响土壤呼吸速率。因此水层深度对土壤呼吸的影响还具有不确定性和复杂性[42]。
研究表明[43],土壤温度和含水量的协同作用能够更好地解释土壤呼吸变化情况,解释能力高于50%,一般比单一因子的解释能力更强,但由于农田生态系统复杂的环境因子,目前水热因子同土壤呼吸之间的关系仍然难以定量描述。
4 结论
通过对川西平原灌区三种不同水旱轮作模式的土壤呼吸特征及其水热影响因素研究,结果表明,土壤呼吸周年累积排放量以GR模式最高,从不同作物生长季来看,稻季>旱季>空闲季。土壤呼吸速率季节动态呈双峰曲线,在6月和9月达到峰值。土壤呼吸速率日动态呈单峰曲线,土壤呼吸速率在12:00—15:00间出现最大值,在2:00—6:00之间出现最低值。三种模式土壤呼吸速率与0—10 cm和10—20 cm土壤温度呈指数正相关且对温度变化敏感,Q10值变化范围在1.15—2.72之间。不同模式间相比,GR模式Q10值最高,土壤呼吸速率与土壤温度敏感性最高。三种模式土壤呼吸与土壤水分没有显著相关性,但稻季与水层高度表现为显著负相关。然而,水热生态因子对水旱轮作土壤呼吸的影响是一个十分复杂的过程,既会直接影响土壤呼吸的排放,也会通过改变土壤微生物区系组成、外源碳的进入量等间接影响土壤呼吸,其作用机制还需要进一步论证研究。总体来看,川西平原灌区水旱轮作栽培管理过程中,应注意控制蒜-稻轮作模式的氮肥投入,稻季合理管理田间水层,从而有效管理土壤呼吸碳排放。
猜你喜欢
农村实用技术(2021年9期)2021-10-30
南方水产科学(2021年3期)2021-06-23
水土保持研究(2019年6期)2019-10-19
吉林农业(2018年15期)2018-08-14
中国水土保持科学(2017年3期)2017-07-05
现代农业科技(2017年5期)2017-04-19
农业与技术(2017年2期)2017-03-16
现代农业科技(2016年21期)2017-03-06
浙江大学学报(工学版)(2016年2期)2016-06-05
天然气勘探与开发(2014年4期)2014-02-28
