
长期氮添加对亚热带森林土壤微生物碳源代谢多样性的影响
2019-11-07 06:43:00鲁显楷张永清莫江明于贵瑞
生态学报 2019年18期
刁 婵,鲁显楷,田 静,张永清,莫江明,于贵瑞
1 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室,北京 100101 2 山西师范大学,地理科学学院,临汾 041000 3 中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广州 510650
大气氮(N)沉降是陆地生态系统重要的N输入过程,对维持生态系统N平衡和生产力至关重要[1]。然而,人类活动加速了N的输入过程,据估计,过去145年(1860—2005),全球N沉降增加了近3倍,并预计到2050年将达到200 Tg N a-1[2]。大量研究证实大气N沉降的频繁输入会引起土壤呼吸、养分矿化与固定、土壤微生物分解等一系列生态过程不同程度的变化[3- 4]。而这些变化加速又改变着土壤微生物多样性以及土壤养分循环[5- 6]。
土壤微生物作为土壤生态系统重要的组成部分,推动着生物地球化学循环与转化,是衡量土壤肥力与健康的关键指标[7- 8],因此微生物多样性对土壤质量及生态系统稳定性具有重要意义。大量研究表明,N沉降会显著影响土壤微生物量和群落结构[9- 14]。目前研究发现土壤微生物量对N沉降具有增加[11]、减少[12- 13]或不变[14]等3种响应。微生物功能多样性信息对于明确不同环境中微生物群落的作用具有重要意义[15]。其中以BIOLOG微孔板碳源利用为基础的定量分析描述了微生物对不同种类碳源的利用多样性,被证明是研究微生物群落代谢功能多样性的一种简单和快速的方法[15]。目前研究发现氮沉降增加会引起土壤微生物对底物的利用模式改变。例如,Johnson等[16]在欧石南丛生的荒地中的研究发现,氮增加造成了微生物对底物的利用率提高了3倍;但在酸性的草地中则相反,土壤微生物对氮增加的响应表现为降低对碳源的利用率。Frey等[12]在美国哈佛森林氮添加实验发现,阔叶林和混交林土壤微生物对底物的利用率与氮增加并没有明显的关系。这些研究的差异性可能和研究区域,施N时间以及各区域的土壤植被环境条件有很大关系[17- 19]。比如研究发现土壤微生物对不同碳源的利用情况对外界环境变化比较敏感,会受到多种生物和非生物因素的影响[15,20- 23]。例如Blagodatskaya等[20]发现pH是影响土壤微生物对碳源的利用活性的重要影响因子。陈法霖等[18]通过对南方红壤丘陵区的研究发现,针-阔混合凋落物比单一针叶凋落物更有利于提高土壤微生物群落的碳源利用能力。亚热带森林作为重要的可再生经济资源,其对涵养水源、保持水土、温室气体预算和维持生物多样性方面具有非常重要的贡献[24]。但是随着经济社会发展水平的提升,过量的N输入显著影响了热带和亚热带森林的生物地球化学循环,比如土壤酸化、水体富营养化、林蓄积量下降以及P元素限制加剧等一系列问题[25- 26]。虽然近年来N沉降对亚热带和热带森林生态系统微生物影响的研究越来越多[24,27- 28],但总体上来说研究的时间尺度比温带森林短得多。比如,Cusack等[27]以Luquillo试验林中的热带雨林和山地森林为例分析了N添加对土壤微生物群落特征的影响。袁颖红等[25]探讨了福建省杉木人工林中土壤微生物功能多样性对N添加响应。因此,加强亚热带森林生态系统长期N输入对土壤微生物功能影响的研究,将有助于帮助我们对N、P养分循环等生态系统关键过程的调控,以及对森林生态系统功能动态变化的预测。
本研究以鼎湖山季风常绿阔叶林(以下简称“季风林”)为研究对象,基于已有的长期野外模拟N沉降试验样地平台,运用Biolog微平板技术研究了长期N添加对土壤微生物碳源代谢多样性变化的规律,同时探讨了微生物碳源代谢多样性与环境因子之间的相关关系。研究结果可为热带森林生态系统应对全球变化提供重要理论依据,同时对于推动森林生态系统可持续发展和加强对其管理和功能调控也具有重要的科学意义。
1 材料与方法
1.1 样地概况与设计
本研究位于广东省中部的鼎湖山自然保护区(112°33′E,23°10′N),属于亚热带季风气候。年均温为21.4 ℃,最冷月(1月)和最热月(7月)的均温分别为12.6 ℃和28.0 ℃。年均降雨量为1927 mm,约有3/4集中在3—8月。气温及降水具有明显的季节性特征。土壤类型为由泥盆纪厚层变质砂岩、砂页岩发育而成的不同深度的氧化土(土壤深度超过60 cm)[29- 30]。季风林为本地典型的地带性植被,其乔木层的主要物种有锥栗(Castanopsischinesis)、荷木(Schimasuperba)、厚壳桂(Cryptocaryachinensis)、黄果厚壳桂(Cryptocaryaconcinna)、华润(Machiluschinesis)等;而灌木层则以柏拉木(Blastuscochinchinensis)、九节(Psychotriarubra)、光叶山黄皮(Randiacanthioides)等可耐阴植物类型占比较高[30- 31]。建立样地之初,各小区植被物种组成不存在显著差异。
该研究区域从1990年以来氮沉降问题日趋严重(>30 kg N ha-1a-1),根据观测,2009—2010年,该区域全年N湿沉降量和干沉降量分别为34.4 kg N hm-2a-1和14.2 kg N hm-2a-1[32]。结合本地氮沉降情况,本研究设计了4个N水平处理[29],分别包括:对照(Control:0 g m-2a-1)、低氮(Low N: 5 g N m-2a-1)、中氮(Mediate N:10 g N m-2a-1)、高氮(High N:15 g N m-2a-1)。自2003年7月开始,以NH4NO3为氮源,每月月初以溶液的形式喷洒到样地地面,全年平均喷施,并在整个研究期间持续进行。根据氮处理水平的需要,将每个样方每次所需要喷施的NH4NO3溶解在20 L水中, 对照(CK)、低氮(LN)、中氮(MN)、高氮(HN)四个处理溶液的浓度分别为0、0.3、0.6和0.9 mol N/L。然后以背式喷雾器人工来回均匀喷洒在林地上,喷洒高度1.5 m左右(为了确保肥料的均匀分布,在每个地块上都有两条路径)。对照样方则喷洒等量的水。
1.2 土壤样品的采集与处理
2015年10月在季风林中每个样方内进行采样(12个样方)。每个样方内利用“五点混合法”用土钻(直径4 cm)采集0—10 cm的土壤,然后4 ℃低温保存运往实验室。将用于测定土壤无机氮和可溶性有机碳(DOC)浸提以及Biolog实验的土壤4 ℃保存,并于一周内完成测定;其余土样风干并用于土壤理化性质的测定。
1.3 试验方法
1.3.1土壤理化性质的测定
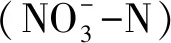
1.3.2土壤微生物碳源利用测定分析
土壤微生物群落碳代谢多样性采用Biolog方法进行[34]:在超净工作台中,称取10 g鲜土置于无菌锥形瓶中,加入90 mL 85%灭菌NaCl溶液,封口后,振荡30 min(250 r/min)。静置10 min后取5mL上清液,将溶液稀释1000倍。最后用8通道移液器将上述稀释液加到Biolog-ECO微平板(Biolog, Hayward, USA)的每个孔中,每孔150 μL。将接种好的Biolog-ECO微平板于25 ℃条件下培养,分别于0、12、24、36、48、60、72、96、120、144、168 h在微孔板读数仪(Biolog, Inc, USA)上读取590nm波长下的吸光值。Biolog-Eco平板测定的每孔颜色平均变化率(average well color development,AWCD),用来表示微生物群落利用单一碳源的能力即微生物的整体代谢活性,计算公式如下[35]:
式中,C为有碳源的每个孔的光密度值,R为对照孔的光密度值,n为碳源的数目,Biolog-Eco板的C源数目为31[36]。
本研究采用培养72 h光密度值分析土壤微生物群落功能多样性,计算公式如下[37]:
Shannon-Wiener多样性指数
丰富度指数S=被利用碳源的总数目(C-R>0.2的孔数)
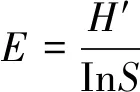
式中,Pi=(C-R)/∑(C-R)表示第i个孔的相对吸光值与整个微平板相对吸光值总和的比值。

图1 不同浓度N处理下土壤微生物群落AWCD值随培养时间的变化Fig.1 Changes of AWCD of soil microorganism under different nitrogen additions with induction timeCK: 对照;LN: 低氮处理; MN: 中氮处理; HN:高氮处理
1.4 数据统计分析
不同处理下土壤微生物功能多样性指数和碳源利用强度的差异用LSD检验确定(SPSS22.0)。在Canoco for Windows 5.0软件中,利用培养96 h的AWCD数据对不同N处理下土壤微生物碳源代谢多样性进行主成分分析(PCA)。用非参数的多元方差分析(PERMANOVA)检验不同N水平处理下土壤微生物碳源代谢多样性的差异。对季风林中环境因子和培养96 h的AWCD数据进行典型对应分析(CCA)和方差分解分析(VPA),探讨环境因子对微生物碳源代谢多样性的影响。其中CCA和VPA通过R语言“vegan”程序包进行。
2 结果
2.1 长期不同N量添加对土壤微生物碳源代谢活性影响
由图1所知,随着培养时间的延长,不同N水平处理下的平均吸光值(AWCD)值均呈现出逐渐增加的变化趋势,并且曲线为平滑的“S”型曲线。0—24 h,土壤微生物的AWCD值缓慢变化;24—96 h AWCD值快速增长,随后增长变缓慢,在96 h后趋于平缓。在整个培养过程中,季风林中AWCD值的大小顺序为:CK>LN>MN ≈ HN。对整个培养过程做重复性方差分析发现:与CK相比,长期MN(P=0.024)和HN(P=0.015)处理显著降低了土壤微生物碳源代谢活性(AWCD)。
2.2 长期不同N量添加对不同碳源利用强度以及多样性影响
Biolog-Eco板中含有31种碳源,包括糖类(7种)、羧酸类(9种)、氨基酸类(6种)、胺类(2种)、聚合物类(4种)、酚酸类(3种)。不同N水平添加处理下土壤微生物群落对同一类型碳源的利用强度存在明显差异。具体表现在:随着N添加水平的提高,微生物对糖类、羧酸、氨基酸、胺类和酚酸类的利用率显著降低;尤其是HN处理最为明显,与CK(对照)相比分别降低了35.3%、48.9%、41.6%、80.2%和32.8%。不同N水平处理没有影响聚合物类的利用。
为了进一步确定N添加对土壤微生物碳源利用率的影响,本文计算了Shannon多样性指数、丰富度指数和均匀度指数(表1)。与CK相比,长期MN和HN处理显著降低了碳源丰富度指数以及Shannon多样性指数。而均匀度指数结果表明,MN和HN显著高于CK。

图2 季风林中不同浓度N处理下土壤微生物碳源利用强度Fig.2 Utilization intensities of carbon sources under different nitrogen additions柱形上方不同小写字母表示不同处理间差异显著(P< 0.05)
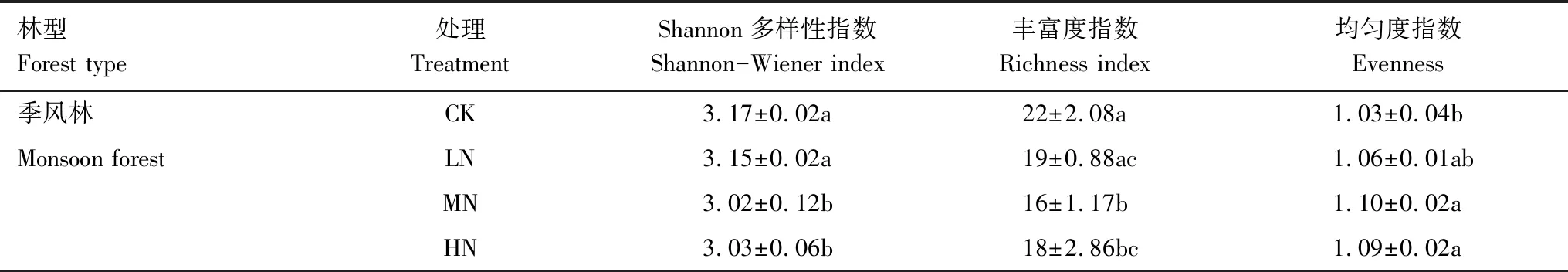
林型Forest type处理TreatmentShannon多样性指数Shannon-Wiener index丰富度指数Richness index均匀度指数Evenness季风林CK3.17±0.02a22±2.08a1.03±0.04bMonsoon forestLN 3.15±0.02a 19±0.88ac 1.06±0.01abMN3.02±0.12b16±1.17b1.10±0.02aHN3.03±0.06b 18±2.86bc1.09±0.02a
同列不同字母表示差异显著(P< 0.05)
2.3 分析长期不同N量添加对土壤微生物碳源利用主成分分析
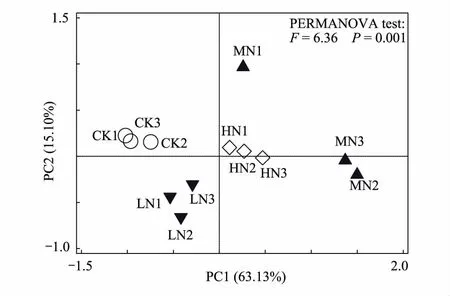
图3 不同浓度N处理下土壤微生物群落对碳源利用多样性的主成分分析Fig.3 Principal components analysis (PCA) of soil microbial carbon source utilization diversity under different nitrogen additions
利用培养96 h的AWCD数据,对31种碳源利用情况进行主成分分析(图3)。主成分1(PC1)和主成分2(PC2)的方差贡献率分别为63.1%和15.1%,累计方差贡献率为78.2%,说明PC1和PC2是微生物群落碳源利用变异的主要来源,可以解释变异的大部分信息。统计检验表明不同N水平添加对土壤微生物群落在碳源利用上有明显的分异(图3,PERMANOVA检验,P=0.001)。
2.4 长期不同N量添加下土壤微生物碳源利用的主要环境影响因子
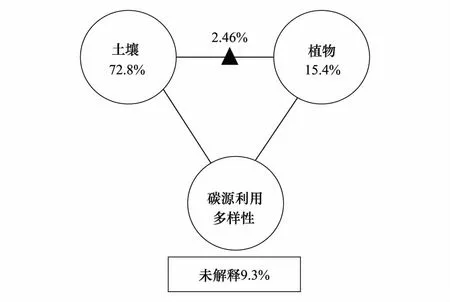
图4 不同浓度N处理下土壤微生物群落碳源利用环境因子影响的分类变异分析Fig.4 Variation partitioning analysis (VPA) of carbon source utilization diversity and environmental factors under different nitrogen additions
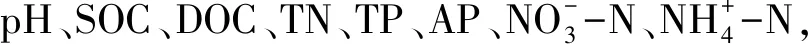
为了更好的明确影响土壤微生物碳源利用的主要显著影响环境因子,将与之相关的10个环境因子进行了典型对应分析(CCA)(图5)。检验结果如下:引起不同N添加下土壤微生物碳源利用显著差异最主要的土壤因子是土壤pH(F=4.78,P=0.009)。植被因子(植被丰富度和凋落物量)与土壤微生物碳源利用差异无显著相关性。

图5 不同浓度N处理下土壤微生物群落碳源利用多样性与环境因子的典型对应分析Fig.5 Canonical correspondence analysis (CCA) of carbon source utilization and environmental factors under different nitrogen additions
3 讨论
本研究发现不同N水平添加显著影响了土壤微生物对不同碳源的利用能力以及碳源利用多样性。一般来说,土壤微生物碳源利用率和AWCD值之间成正比例关系,即AWCD可反映土壤微生物活性对碳源利用能力或利用强度[24,38]。本研究中,季风林中土壤微生物碳源利用强度随N添加量的增加而显著降低,和Compton等[39]在Harvard森林长期氮输入实验结果一致。我们还发现糖类、羧酸、氨基酸、胺类和酚酸类碳源的利用随着施N水平的提高显著下降(图2)。长期高剂量N添加显著降低土壤微生物碳源利用率的原因可能在于:一方面,长期施用N肥,不仅会降低微生物生物量[40- 41],而且对土壤微生物活性产生一定的负面影响[42],进而影响土壤微生物对底物的利用率;同时季风林样地为相对富N环境[32],土壤中N元素基本上可能已经达到了饱和的临界值,如果继续施N肥,也会对微生物对碳源利用率产生一定的抑制作用[43]。Deforest等[42]在北方硬叶阔叶林氮沉降样地中的试验以及刘蔚秋等[44]在南亚热带林内生境中微生物底物利用能力影响的研究中也得出了类似的结论。但是也有研究发现施用N肥会提高微生物碳源利用多样性或者无影响,比如Grayston等[45]在英国高山草地中发现,N增加提高了土壤微生物的碳源利用效率。Zhou等[14]则在对位于中国西北部的古尔班通古特沙漠的研究表明,土壤微生物对底物的利用率与N增加并没有明显关系。这些研究的差异可能主要与研究区域、施氮时间、研究时期等有关。
多样性指数从本质上来说,一方面可以反映土壤微生物群落功能多样性,另一方面也是评价土壤微生物群落利用碳源程度的重要指标,该值和微生物群落的多样性、碳源利用效率之间有着正相关关系[46]。不同的碳源利用多样性指数又可真实的反映土壤微生物群落功能多样性的不同侧面。本研究表明施N对土壤微生物的功能活性产生了显著的影响,主要体现在Shannon指数和丰富度指数均随着N浓度的增加均表现出降低的趋势(表2)。部分研究也发现LN处理在不同程度上增加了均匀度和Shannon指数,而HN处理下则出现降低[22,47]。土壤微生物对土壤生态系统环境的改变反映非常的敏感[48]。不同浓度N添加处理可以通过直接或间接影响土壤理化性质、物质转换以及营养物质的有效性,使土壤微生物生存的微环境发生改变,进而导致土壤微生物结构及其功能多样性发生变化。MN和HN处理下土壤微生物功能多样性显著低于对照处理可能是多种环境因子共同作用的结果,其中主要包括土壤因子和气候因子的多重作用以及两者之间的交互影响(图4、5)。
土壤pH(P<0.01)对土壤微生物碳源代谢产生极其显著的影响的主要原因可能是施N量的增加会直接导致土壤pH的下降,进而对土壤中微生物群落的群落结构和活性产生显著影响。刘蔚秋等[44]通过对南亚热带林内生境中土壤微生物对模拟大气氮沉降的响应研究,以及Smolander等[49]、Aerts等[50]和Wallander等[51]的研究表明,持续的N增加使土壤含N量处于过饱和状态,由此引起的土壤酸化、板结和毒效应可能是微生物功能改变、活性降低的重要原因。其他研究中曾发现类似的研究现象[52-54]。植物因素对土壤微生物碳源利用效率变异的解释程度较小,这可能与该样地林龄较长而处于缓慢生长期等因素有关。
4 结论
长期不同N量添加显著降低了土壤微生物碳源代谢活性,同时改变了土壤微生物对底物(不同碳源)的利用模式,进而引起微生物碳源利用多样性发生相应的改变。土壤pH因子是长期不同N水平处理下微生物碳源利用变异的主要影响因素。该研究结果可为热带森林生态系统生物多样性和生态系统功能改变对全球变化响应提供了理论依据和数据支撑;同时对于推动森林生态系统可持续发展和加强对其管理和功能调控也具有重要的科学意义。
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05 07:20:20
中等数学(2022年2期)2022-06-05 07:10:50
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
昆钢科技(2021年6期)2021-03-09 06:10:20
小学生学习指导(低年级)(2020年6期)2020-07-25 02:31:36
小学生学习指导(低年级)(2018年9期)2018-09-26 05:59:44
疯狂英语·新读写(2018年2期)2018-09-07 09:32:10
电源技术(2016年9期)2016-02-27 09:05:25
水生生物学报(2015年1期)2015-02-28 16:01:05
