
播期、品种对沙地紫花苜蓿抗氧化特性与越冬率的影响
2019-11-07 10:41田永雷张玉霞王显国候文慧王月林孙明雪杜晓艳
草地学报 2019年5期
田永雷,张玉霞,王显国,候文慧,王月林,孙明雪,杜晓艳
(1.内蒙古民族大学农学院,内蒙古 通辽 028041; 2.内蒙古自治区农牧业科学院草原研究所,内蒙古 呼和浩特 010031;3.中国农业大学动物科技学院,北京 100193; 4.内蒙古自治区草原工作站,内蒙古 呼和浩特 010017)
紫花苜蓿(MedicagosalivaL.)是豆科苜蓿属多年生牧草,有“牧草之王”的誉称[1]。由于紫花苜蓿具有高产、优质、保持水土、改良土壤、抗旱性较强等优点,是目前我国广泛种植的优质牧草[2],近些年,在北方干旱地区种植面积不断增大,然而在北方干旱地区,尤其是科尔沁沙地由于风沙大,降雨少,春季温度低等因素[3],加之种植方式不当,导致紫花苜蓿越冬不稳,严重限制紫花苜蓿持续稳定生产[4-6]。目前我国关于苜蓿播期的研究主要是针对播期对苜蓿的生产性能和形态特征以及越冬率的研究[7-9],关于紫花苜蓿的抗氧化机理研究多集中在低温胁迫下的紫花苜蓿根颈和叶片生理变化[10-15],有关不同播期下低温驯化期紫花苜蓿根颈抗氧化变化的报道较少[16-18],尤其是在科尔沁沙地地区研究未见报道。为此,在科尔沁沙地,开展紫花苜蓿不同播种时期试验,探讨越冬前期紫花苜蓿根颈的抗氧化特性变化及其与越冬率和播期之间的关系,阐述播期影响科尔沁沙地紫花苜蓿越冬的抗氧化生理机理,提出适宜的播种日期范围,为科尔沁沙地紫花苜蓿适宜时期种植和抗寒性研究提供理论依据。
1 材料与方法
1.1 试验区自然概况
试验地位于内蒙古自治区赤峰市阿鲁科尔沁旗惠农草业有限公司牧草生产基地,地理位置为北纬42°30′50.4″,东经115°27′34.2″,海拔322 m。该地区为温带半干旱大陆性气候,年平均气温5.5℃,≥10℃的积温为3 000~3 200℃,无霜期140~150 d,年平均降水量约375 mm,年平均风速3.0~4.4 m·s-1,土壤为沙壤土。试验地土壤营养成分碱解氮含量38.8 mg·kg-1,速效磷含量2.1 mg.kg-1,速效钾含量103 mg·kg-1,有机质含量7.28 mg·kg-1,PH值7.26。
1.2 试验设计
试验采用2因素随机区组试验设计。选择骑士T、公农1号、擎天柱3个紫花苜蓿品种,于2017年7月1日至8月15日,每5天1个播种时期,共10个播种时期。小区面积4 m×6 m=24 m2,4次重复,共120个小区。于2017年10月11日和11月11日进行苜蓿根系取样,测定根颈的超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化物酶(Peroxidase,POD)、过氧化氢酶(Catalase,CAT)活性和丙二醛(Malondialdehyde,MDA)含量,2018年4月20日调查越冬率。
1.3 测定指标及方法
丙二醛含量用硫代巴比妥酸法测定[19];SOD酶活性用氮蓝四唑法测定[19];CAT酶活性用紫外吸收法测定[19];POD酶活性用愈创木酚法[20]测定;越冬率=2018年返青存活株数/2017年冬前存活株数×100%[21]。
1.4 数据分析
试验数据用WPS软件处理、做图和制作表格,SPSS 17.0软件进行方差显著性分析,用Duncan法进行多重比较,显著水平0.05。
2 结果与分析
2.1 不同播期下紫花苜蓿的越冬率变化
由图1可知,随着播期的延迟,紫花苜蓿的越冬率逐渐降低,骑士T、公农1号、擎天柱3个紫花苜蓿品种的越冬率均在7月16日播期开始显著降低(P<0.05),不同品种之间越冬率比较公农1号显著高于骑士T和擎天柱(P<0.05)。
2.2 不同播期下紫花苜蓿根颈的SOD酶活性变化
10月11日测定7月1日播种的骑士T根颈的SOD酶活性最高(见表1),显著高于7月11日以后的播期(P<0.05);7月6日播种的公农1号根颈的SOD酶活性最高,与其它播期无显著差异;7月1日播种的擎天柱根颈的SOD酶活性显著高于7月21日以后播期(P<0.05);3个紫花苜蓿品种间SOD酶活性无显著差异。11月11日测定的SOD酶活性均较10月11日测定的SOD酶活性明显降低,其中7月1日播种的骑士T和公农1号根颈的SOD酶活性最高,7月21日播种的擎天柱根颈的SOD酶活性最高,但不同播期间SOD酶活性均无显著差异,3个紫花苜蓿品种间SOD酶活性为擎天柱>公农1号>骑士T,且擎天柱显著高于骑士T(P<0.05)。
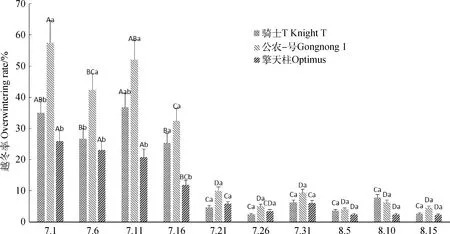
图1 不同播种期下紫花苜蓿的越冬率

测定时间measuringdate播种时间sowingdates品种Varieties骑士TKnightT公农1号Gongnong1擎天柱OPtimus10月11日11thOctober01/07584.55±97.91Aa482.25±33.22Aa534.34±16.22Aa06/07476.35±26.84ABa511.38±35.34Aa512.56±76.35Aa11/07493.69±104.25ABa496.63±31.51Aa526.47±15.63Aa16/07400.25±113.41Ba486.85±49.76Aa494.38±26.27Aa21/07448.69±125.75ABa441.69±29.28ABa508.26±28.76Aa26/07495.75±51.12ABa479.57±52.87Aab395.94±17.67Cb31/07415.58±42.53Ba361.78±68.93Ba473.17±48.49ABa05/08360.85±41.11Ba426.83±49.97ABa406.82±63.52BCa10/08468.96±93.92ABa456.77±67.16Aa368.96±57.83Ca15/08391.74±54.93Bb473.48±29.16Aa528.63±27.37Aa11月11日11thNovember01/07272.85±32.87Ab413.24±64.37Aa422.25±89.33Aa06/07242.98±29.01Ab349.89±61.65ABab430.36±111.34Aa11/07247.96±48.03Ab388.75±71.27ABa439.85±66.25Aa16/07250.25±57.05Ab334.45±43.85ABab437.46±110.27Aa21/07230.78±57.53ABb354.67±48.63ABab446.52±86.07Aa26/07233.96±50.12ABb318.25±24.64ABab405.14±28.32Aa31/07209.47±65.37Bb327.14±59.37ABab362.17±47.96Aa05/08247.56±22.34Ab360.77±21.56ABa333.18±58.57Aa10/08204.52±40.66Bb302.26±41.45ABa351.29±59.98Aa15/08233.58±68.15ABb276.89±17.07Bab342.34±94.21Aa
2.3 不同播期下紫花苜蓿根颈的POD酶活性变化
10月11日测定7月1日播种的骑士T根颈的POD酶活性最高(见表2),显著高于除7月16日、7月31日和8月15日播期以外其它播期(P<0.05);8月15日播种的公农1号根颈POD酶活性最高,但不同播期之间无显著差异;7月1日播种的擎天柱根颈的POD酶活性最高,显著高于7月11日以后播期(P<0.05);3个紫花苜蓿品种间无显著差异。11月11日测定7月16日播种的骑士T的POD酶活性最高,与8月5日、8月10日、8月15日无显著差异,显著高于其它播期(P<0.05);公农1号在7月11日播期时POD酶活性最高,显著高于7月31日以后播期(P<0.05);擎天柱在8月5日播期时POD酶活性最高,不同播期POD酶活性均无显著差异;3个紫花苜蓿品种之间POD酶活性比较骑士T高于公农1号和擎天柱。

表2 播种时期对POD酶活性的影响
2.4 不同播期下紫花苜蓿根颈的CAT酶活性变化
10月11日测定7月6日播种的骑士T根颈的CAT酶活性最高(见表3),显著高于除7月1日以外的其它播期(P<0.05);8月10日播种的公农1号根颈的CAT酶活性最高,显著高于8月15日播期(P<0.05),与其它播期无显著差异;7月11日播种的擎天柱根颈的CAT酶活性最高,显著高于其它播期(P<0.05);3个紫花苜蓿品种间CAT酶活性比较骑士T低于公农1号和擎天柱。11月11日测定8月15日播种的骑士T根颈的CAT酶活性最高,不同播期间CAT酶活性无显著差异;8月5日播种的公农1号根颈的CAT酶活性最高,不同播期间无显著差异;7月21日播种的擎天柱根颈的CAT酶活性最高,显著高于7月6日和7月11日(P<0.05),与其它播期无显著差异;3个紫花苜蓿品种间CAT酶活性比较骑士T低于公农1号和擎天柱。
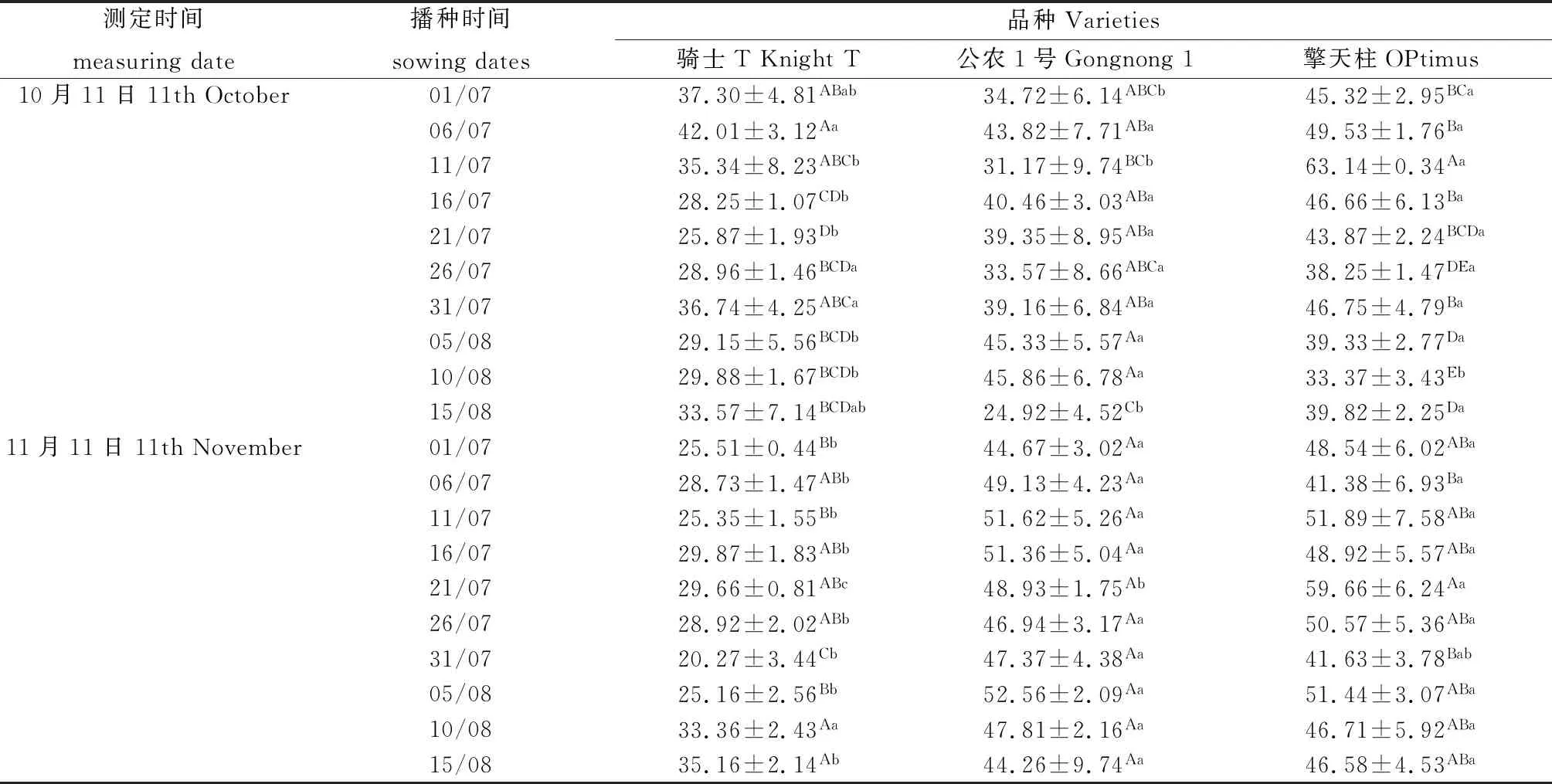
表3 播种时期对紫花苜蓿根颈CAT酶活性的影响
2.5 不同播期下紫花苜蓿根颈的MDA含量变化
10月11日测定7月31日播种的骑士T根颈的MDA含量最高(见表4),显著高于除7月16日以外的其它播期(P<0.05);8月5日播种的公农1号根颈的MDA含量显著高于其它播期(P<0.05);在8月5日播种的擎天柱紫花苜蓿品种根颈MDA含量最高,显著高于除7月16日以外的其它播期(P<0.05);3个紫花苜蓿品种间的MDA含量比较擎天柱小于骑士T和公农1号。11月11日测定7月31日播种的骑士T根颈的MDA含量显著高于7月26日以前的播期(P<0.05);8月10日播种的公农1号的MDA含量显著高于7月1日、7月16日和7月26日播期(P<0.05);8月15日播种的擎天柱的MDA含量显著高于7月1日和7月11日播期(P<0.05),与其它播期差异不显著;3个紫花苜蓿品种间的MDA含量比较骑士T高于公农1号和擎天柱。
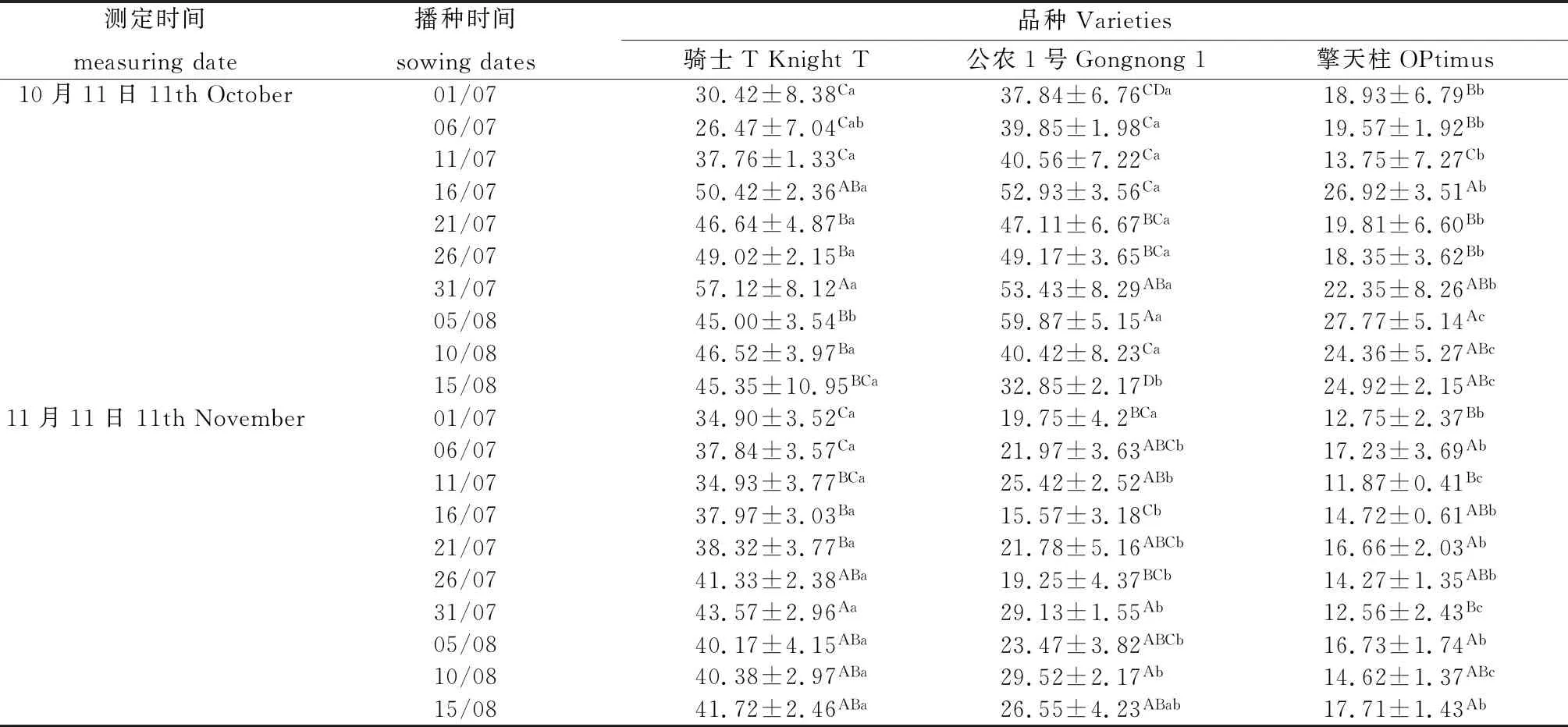
表4 播种时期对MDA含量的影响
2.6 紫花苜蓿抗氧化生理指标与越冬率的相关性分析
由表5可知,10月11日紫花苜蓿根颈的MDA含量与SOD,POD酶活性均呈显著负相关,与CAT酶活性呈极显著负相关;11月11日的紫花苜蓿根颈的MDA含量POD酶活性呈显著负相关,与SOD和CAT酶活性呈极显著负相关,由此说明SOD,POD,CAT3种抗氧化酶是清除低温胁迫下活性氧自由基的关键酶。越冬率与10月11日紫花苜蓿根颈的SOD,POD,CAT酶活性呈正相关,且与SOD酶活性达到显著水平;越冬率与11月11日紫花苜蓿根颈的SOD,POD酶活性呈正相关,且与POD酶活性达到显著水平;越冬率与10月11日和11月11日紫花苜蓿根颈的MDA含量均呈负相关,但没有达到显著水平;越冬率与播期呈极显著负相关。由此说明,播种时期通过影响越冬前期紫花苜蓿根颈的抗氧化酶活性,进一步影响沙地紫花苜蓿的越冬性。

表5 紫花苜蓿抗寒生理指标与越冬率的相关性分析
注:**:在.01 水平(双侧)上显著相关;*:在 0.05 水平(双侧)上显著相关
Note:**:Significant correlation at the 0.01 level (bilateral),*:Significant correlation at the 0.05 level (bilateral)
3 讨论
国内外关于紫花苜蓿抗寒性与根系抗氧化酶活性关系的研究资料较多,研究表明[22-24],紫花苜蓿根系中保护酶(过氧化物酶、超氧化物歧化酶、过氧化氢酶)活性与其抗寒性密切相关[25]。申晓慧[24]研究表明,SOD酶活性与越冬率成正比,本研究中10月11日和11月11日苜蓿根颈中的SOD酶活性与越冬率均呈正相关关系,与其结果一致;寇建村[23]等研究表明,不同紫花苜蓿品种间SOD酶活性存在显著差异,本研究与其结果一致,11月11日测定骑士T紫花苜蓿根颈中SOD活性明显小于公农1号和擎天柱苜蓿;马周文等[26]表明低温处理下SOD酶活性无显著差异,其解释为SOD酶活性对低温胁迫不敏感,本研究与其结果不同,本研究中紫花苜蓿根颈SOD酶活性11月份较10月份下降,可以解释为紫花苜蓿感受寒冷刺激后产生应激能力,当紫花苜蓿获得抗寒能力后,SOD酶活性逐渐下降[27]。
Duke[28]研究表明,抗寒性强的紫花苜蓿品种可以通过提高POD酶活性来提高自身的抗寒性,从而减少低温胁迫对植物的伤害;杨秀娟[29]、申晓慧[24]等结果表明,紫花苜蓿根颈中的POD酶活性与越冬率显著正相关,本研究与其结果一致,越冬率与11月11日测定苜蓿根颈POD酶活性呈显著正相关关系,但与10月11日测定紫花苜蓿根颈中POD酶活性相关性未达到显著水平。朱爱民等[30]研究表明,紫花苜蓿感受低温胁迫后可能会启动CAT保护机制,减少超氧阴离子的伤害,本研究亦能佐证此观点,相关性分析表明10月11日和11月11日测定苜蓿根颈中的CAT活性与丙二醛含量呈极显著的相关关系。
丙二醛是膜质过氧化的最终分解产物,诸多研究表明,丙二醛可以作为衡量植物抗逆性强弱的重要指标。如:陶雅等[12]研究表明,不同品种紫花苜蓿根系在低温初期均有大量丙二醛的积累,随后丙二醛含量会降低,温度升高后,丙二醛含量逐渐增加;申晓慧[31]测定了紫花苜蓿根系整个越冬期生理指标的变化,表明紫花苜蓿根中丙二醛的含量随温度的变化呈现先升高后降低再升高的趋势,与越冬率成反比。本研究表明,紫花苜蓿根颈中丙二醛含量与越冬率呈负相关关系,这与申晓慧的研究结果一致。
随着播期的推迟,紫花苜蓿越冬率明显下降[19];播期较晚可以提高紫花苜蓿出苗率,但播种太晚则不利于提高越冬率[32];播期越早越有利于紫花苜蓿越冬[33];本研究表明,播期与越冬率呈极显著负相关关系,说明在科尔沁地区,播期越晚,紫花苜蓿越冬率越低。
4 结论
随着播期的延迟,紫花苜蓿的越冬率呈下降趋势,在7月21日播种较前期播种下降显著;越冬率与冻融交错期紫花苜蓿根颈部的SOD酶活性和封冻期紫花苜蓿根颈部的POD酶活性呈显著正相关关系;2个测定时期紫花苜蓿根颈部MDA含量与SOD,POD,CAT酶活性均呈显著或极显著负相关关系;在阿鲁科尔沁地区,7月21日之前播种可有效提高紫花苜蓿越冬率。
猜你喜欢
农业工程学报(2022年5期)2022-06-22
当代工人(2020年7期)2020-05-20
西藏农业科技(2019年1期)2019-07-25
学生天地·小学中高年级(2018年8期)2018-10-11
学生天地(2018年23期)2018-09-11
畜牧兽医杂志(2018年2期)2018-06-15
江苏农业科学(2016年8期)2017-02-15
玩具世界(2017年8期)2017-02-06
现代农业科技(2016年20期)2016-12-20
西藏科技(2016年9期)2016-09-26
