
青春期双酚A暴露对雌鼠情绪和社会行为的影响
2019-11-04 11:34殷张昕高彤彤庄紫薇钟晓宇李季穗王文波姜科声胡一中徐晓虹
浙江师范大学学报(自然科学版) 2019年4期
殷张昕, 高彤彤, 庄紫薇, 钟晓宇, 李季穗,王文波, 姜科声, 胡一中, 徐晓虹
(浙江师范大学 化学与生命科学学院,浙江 金华 321004)
0 引 言
BPA是酚类化合物中具有代表性的一类环境内分泌干扰物,因其可以改进塑料制品的可塑性和拉伸性而大量应用于生产碳酸聚酯、环氧树脂类食品饮料包装材料、医疗用品和日常用品,导致人们和野生动物的终生BPA暴露风险.2008年美国健康食品安全检查报告调查2 517人中有92.6%的尿液检出BPA,儿童和青少年体内BPA含量更高[1].BPA的化学结构类似己烯雌酚,可与ER结合而表现出弱雌激素活性,从而干扰内源性激素对机体生理活动的调节,对人类和野生动物的生殖和发育产生危害[1-2].据此,欧美制定了人群BPA的最低容许摄入量为50 μg·kg-1·d-1.然而,近十多年的研究发现,低剂量BPA(<50 μg·kg-1·d-1)可影响人和动物的脑和行为发育[3-5].出生早期BPA暴露可引起雌性大鼠青春期提前[6],妊娠期BPA暴露加重青春期仔鼠的焦虑和抑郁样状态[7],并消除焦虑和抑郁行为的性别差异[8-9],还可影响脑内神经元突触发育[10].
青春期是脑发育和社会认知形成的关键时期,此时开始发育的性腺分泌性激素,对社会行为的神经环路成熟和功能起激活作用,对社会行为的形成及其性别分化非常重要[11].社会玩耍是最早发生于非母亲指向的幼年至青春期同种个体间的社会行为,有利于个体社会技能和认知功能发育.早期社会隔离和玩耍剥夺会减少青少年社会动机,使成年后社会互作减少,攻击行为增加,并影响性行为[12].社会行为异常还是早期精神发育障碍的主要症状,自闭症和早发性精神分裂症患者在儿童至青春期即表现出社会玩耍和社会互作减少[13].青春期性激素对社会行为神经环路成熟和功能激活的重要作用,使青春期的脑对环境内分泌干扰物非常敏感[14].流行病学研究发现,出生前后BPA暴露导致儿童攻击、注意缺陷、多动障碍、焦虑和抑郁等多种精神和情绪异常[15].因此,推测青春期暴露于BPA很有可能影响雌性动物的社会行为,但目前国内外还没有相关报道.本文探讨了青春期BPA暴露对雌鼠情绪和社会行为的影响,并分析是否与DA递质系统活动的改变有关.
1 材料和方法
1.1 实验动物、模型制备与分组
3周龄ICR雌性小鼠购于浙江省医学科学院动物中心,常规饲养适应1周后,参考人群日常暴露水平,设3个BPA暴露组:0.04,0.40,4.00 mg·kg-1·d-1(根据体表面积药理剂量换算相当于人体暴露水平:3.24,32.4,324 μg·kg-1·d-1)和溶媒对照组(油).每天灌胃染毒BPA连续3周,制备青春期BPA暴露模型.每组小鼠分2笼饲养.在8周龄(青年期)时,每组随机从各笼中取12只小鼠进行行为检测(n=12),另取未进行行为测试的8只小鼠眼眶取血,酶联免疫分析法检测小鼠脑和血清雌二醇(estradiol,E2)水平(n=8),取6只小鼠提取脑纹状体组织用于蛋白表达检测(n=6),最后分离子宫称重,计算子宫系数(子宫质量/体质量).
1.2 行为测试
行为测试按以下顺序进行.整个过程用小动物行为分析系统(德国TSE 系统股份有限公司)记录.每一行为结束后,隔天进行下一个行为检测.每只小鼠测试后用75%乙醇擦拭模型,防止气味影响下一个动物的行为.所有行为由小动物行为分析系统自动监测记录,并由经过训练的人员采用双盲法采集和分析数据.
采用旷场行为[16]记录小鼠5 min内的跑动穿越格数、中央区停留时间、站立和理毛次数(n=12).其中,跑动格数反映动物在新异环境中的自发活动性,站立次数反映动物的探究意愿,中央区停留时间反映焦虑状态.
采用社会玩耍行为[16]记录15 min内2只相同年龄和处理的陌生小鼠互相爬跨、追逐、打斗、理毛和嗅探等玩耍指标,其中互相理毛和嗅探作为社会探索(n=12)[17].
采用三室模型[16]记录5 min内待测小鼠分别在含陌生鼠圆筒或含空圆筒的隔间所停留时间,以及对陌生鼠或空圆筒的嗅探时间来检测小鼠的社会交互能力(n=12).测试小鼠在含陌生鼠隔间停留或嗅探陌生鼠的时间越长,社会交互能力越强[18].
采用高架十字迷宫[16]记录5 min内动物在开放臂的停留时间来评价焦虑情绪,中央区探头次数反映动物的探究意愿(n=12).
采用强迫游泳模型[16]检测动物的习得性无助而评价其抑郁状态[19].记录小鼠从游泳开始至出现第1次静止不动的时间记为潜伏期;记录6 min强迫游泳期的后4 min内小鼠静止不动的总时间.静止不动的潜伏期越短或静止不动总时间越长表示抑郁越严重(n=12).
1.3 Western Blot检测纹状体组织ERβ,DAT和 DR1蛋白表达水平
小鼠(8周龄)断颈处死后取脑,4 ℃分离双侧纹状体组织,制备蛋白样品分装,-20 ℃保存备用(n=6).取10~20 μL蛋白样品SDS-PAGE凝胶电泳,将蛋白湿转至硝酸纤维素膜(NC)上,封闭,分别加相应一抗(β-actin(1∶1 000),ER-β(1∶400),DAT(1∶500),DR1(1∶600))孵育,漂洗,再将NC膜与辣根过氧化物酶标记的羊抗兔二抗孵育,漂洗,用免疫化学发光试剂显色,X片曝光后显影、扫描,Quantity One软件半定量分析蛋白条带光密度.
1.4 统计学处理
所有数据用平均值±标准误差(M±SE)表示,采用SPSS 18.0统计软件通过单因素方差分析不同剂量BPA暴露的作用,进行Post-hoc 测试后LSD检验,P<0.05表示具有显著性差异,P<0.01表示具有极显著性差异.
2 结果与分析
2.1 青春期BPA暴露对雌鼠生长发育的影响
青春期0.04,0.40,4.00 mg·kg-1·d-1BPA暴露没有明显改变青年雌鼠体质量和子宫系数,提示青春期低剂量BPA暴露对小鼠的生长和生殖器官发育没有明显影响(见表1).
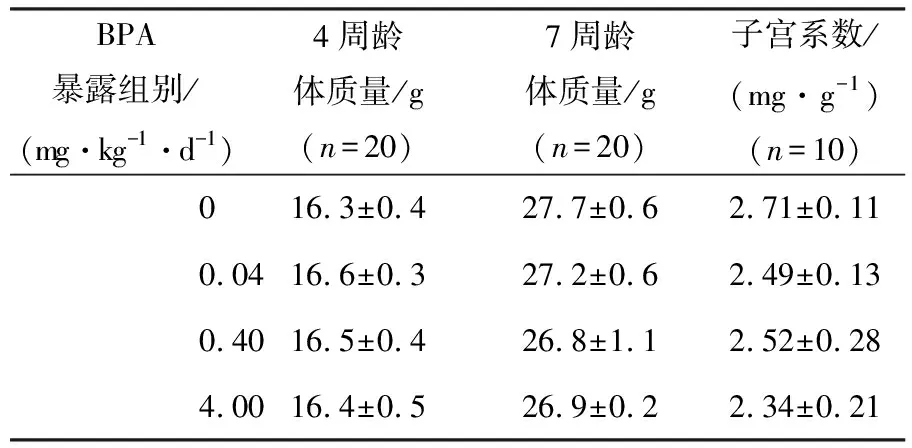
表1 BPA对雌鼠体质量和子宫系数的影响
2.2 青春期BPA暴露对雌鼠旷场行为的影响
4.00 mg·kg-1·d-1BPA显著减少小鼠在旷场中悬空站立次数(P<0.05),0.40,4.00 mg·kg-1·d-1BPA显著减少小鼠在中央区的停留时间(P<0.05),提示青春期BPA暴露显著降低小鼠的探究意愿,并提高其焦虑状态(见表2).
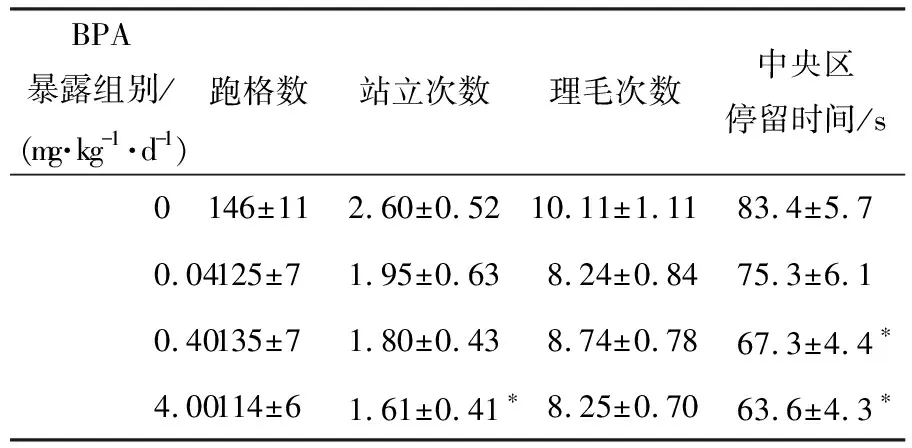
表2 青年雌鼠的旷场行为(n=12)
注:与对照组相比,*表示具有显著性差异(P<0.05).
2.3 青春期BPA暴露对雌鼠高架十字迷宫和强迫游泳行为的影响
高架十字迷宫行为测试结果显示,青春期0.40和4.00 mg·kg-1·d-1BPA暴露显著缩短小鼠在开放臂的停留时间(P<0.01,P<0.05),延长其在封闭臂的停留时间(P<0.05),0.04,0.40,4.00 mg·kg-1·d-1BPA显著减少小鼠中央区探头次数(均为P<0.05)(见表3).强迫游泳测试结果显示,0.40和4.00 mg·kg-1·d-1BPA显著了缩短小鼠强迫游泳时第1次静止不动的潜伏期(P<0.05)和增加了静止不动的总时间(P<0.05)(见图1a~b),提示青春期BPA暴露加重雌鼠的焦虑和抑郁样情绪,降低了其探究的意愿.
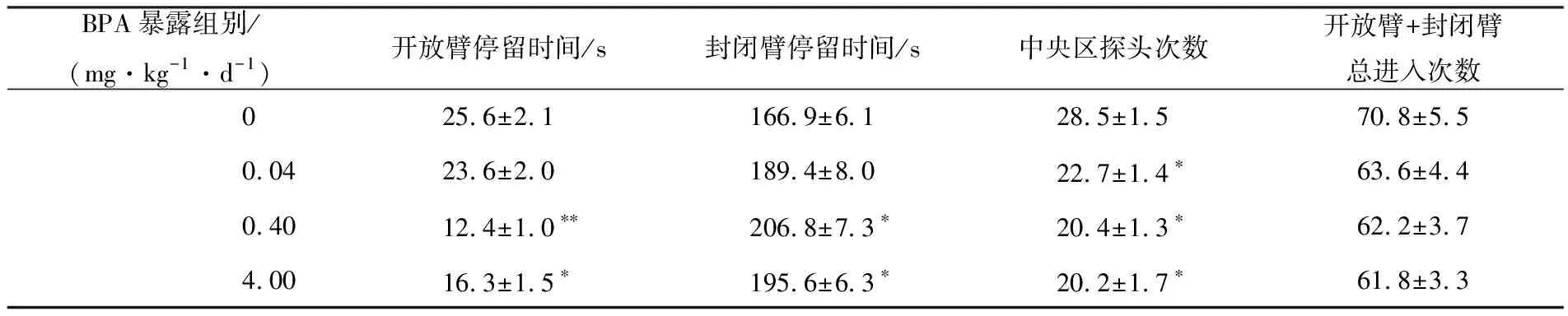
表3 青年雌鼠的高架十字迷宫行为(n=12)
注:与对照组相比,*表示具有显著性差异(P<0.05);**表示具有极显著性差异(P<0.01).
2.4 青春期BPA暴露对雌鼠社会玩耍和社会交互能力的影响
社会玩耍行为测试中,青春期0.40和4.00 mg·kg-1·d-1BPA暴露显著减少小鼠的社会玩耍时间(P<0.05),0.04,0.40,4.00 mg·kg-1·d-1BPA显著减少社会探索时间(P<0.05,P<0.01)和探索频率(4.00 mg·kg-1·d-1BPA,P<0.01);三室模型检测结果显示,与对照组相比,BPA暴露虽然没有明显改变小鼠对陌生鼠或空圆筒的嗅探时间,但0.40和4.00 mg·kg-1·d-1BPA显著降低了小鼠嗅探陌生鼠和嗅探空圆筒的时间比(P<0.05),提示青春期BPA暴露不仅显著降低小鼠的社会玩耍能力,还减弱其社会探究意愿和社会交互能力(见表4).
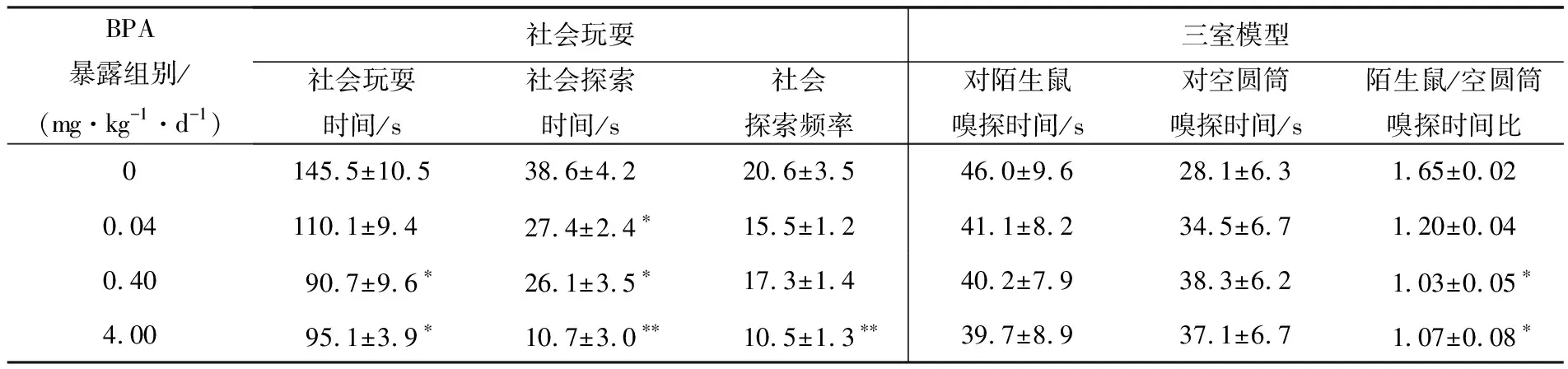
表4 青年雌鼠的社会玩耍和社会交互能力(n=12)
注:与对照组相比,*表示具有显著性差异(P<0.05);**表示具有极显著性差异(P<0.01).
2.5 青春期BPA暴露对成年雌鼠纹状体DAT和DR1表达的影响
鉴于纹状体的DA递质系统在情绪和社会行为中的关键作用,实验进一步分析了纹状体DA转运体(DAT)和DA的D1受体(DR1)蛋白表达水平.研究结果显示,0.40和4.00 mg·kg-1·d-1BPA显著上调DA转运体(DAT)的表达水平(P<0.05,P<0.01),0.04,0.40,4.00 mg·kg-1·d-1BPA显著上调DR1的表达水平(P<0.05,P<0.01)(见图2a~b).
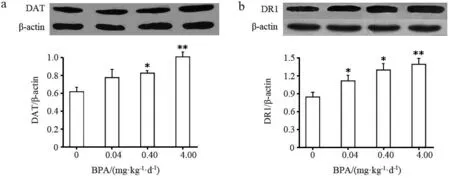
与对照组相比,*表示具有显著性差异(P<0.05);**表示具有极显著性差异(P<0.01)
2.6 青春期BPA暴露对青年雌鼠脑和血清雌激素水平以及纹状体ERβ表达的影响
由于雌激素通过脑内雌激素受体参与调节情绪和社会行为,而纹状体又与情绪和社会行为密切相关,实验检测了脑和血清的雌二醇水平,并用Western Blot分析了纹状体ERβ蛋白表达.研究结果显示,青春期0.04,0.40,4.00 mg·kg-1·d-1BPA暴露没有明显改变青年雌鼠脑和血清雌二醇水平(见图3a~b),但0.40和4.00 mg·kg-1·d-1BPA显著下调纹状体ERβ表达水平(P<0.01)(见图3c).
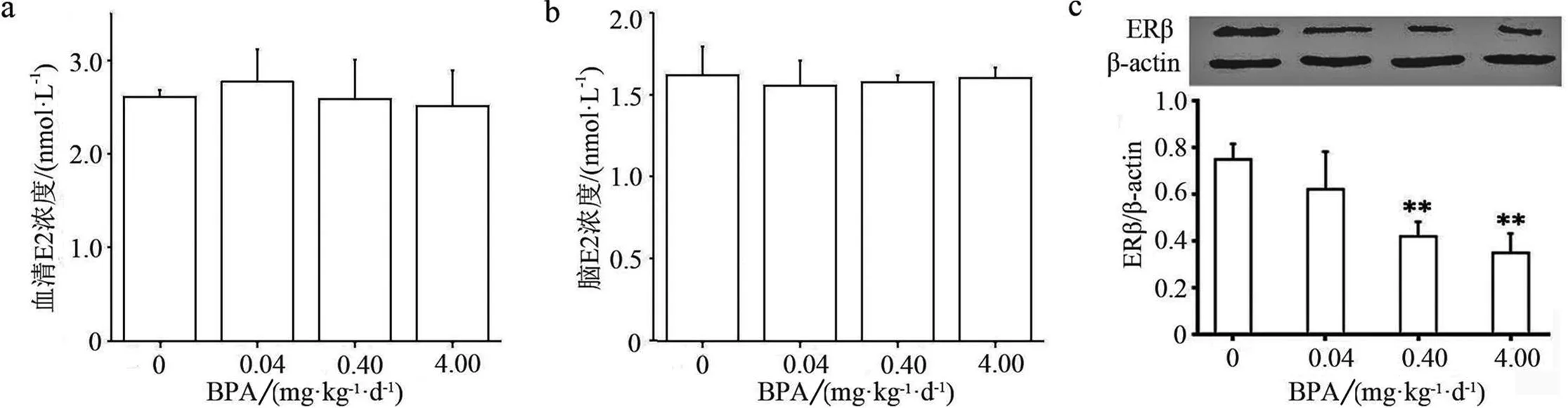
与对照组相比,*表示具有显著性差异(P<0.05);**表示具有极显著性差异(P<0.01)
3 讨 论
青春期作为儿童到成年的一个过渡期,性腺开始发育,此时分泌的性激素对脑和行为发育至关重要.有人认为在出生前后,脑内性激素主要调控行为相关神经环路的形成;而在青春期,性激素对这些环路的成熟和功能起激活作用,是脑和行为发育的第2个关键期[11].因此,青春期的脑对环境内分泌干扰物很敏感.在动物的一生中,最早非母亲指向的社会行为发生在幼年至青春期个体间的玩耍.社会玩耍从社会探究开始,表现为2只陌生小鼠通过相互嗅探熟悉对方,进行追逐、打斗、理毛和爬跨等玩耍活动,是社会交互的一种形式,也是个体融入社会的开始[17-18],对动物的生存和成年后的繁殖至关重要.
近年来动物研究发现,宫内暴露于BPA的青春期猕猴和小鼠与同性同伴的社会互动减少,该行为的性别差异被扰乱[20-21],围生期BPA暴露减少青春期雌鼠的社会探究行为[22],但目前还不了解青春期BPA暴露对社会行为的影响,其机制更不清楚.实验发现,青春期BPA暴露显著降低雌鼠的社会玩耍能力,并减少其相互嗅探和相互接触等社会探索行为.同时在三室模型的陌生同伴和空圆筒的选择中,一般动物更愿意与陌生同伴接触,即具有一定的社会探究意愿.但青春期BPA暴露的雌鼠成年后接触陌生鼠的时间有所缩短,而对空圆筒的接触时间延长,明显降低陌生鼠/空圆筒的嗅探时间比,表明青春期BPA暴露降低雌鼠成年后社会玩耍及社会探索意愿,减弱其社会互作能力.
情绪是影响动物社会行为的一个因素,焦虑和抑郁状态的动物往往表现出较差的社会玩耍和社会交互能力.流行病学研究发现,妊娠期母体尿液高BPA水平与3~12岁儿童表现出的高焦虑和抑郁情绪、及多动症相关[23-24].出生前后BPA暴露的儿童还表现出攻击行为、自闭症等多种精神和情绪异常现象[15].本研究结果显示,0.40和4.00 mg·kg-1·d-1的BPA加重雌鼠在高架十字迷宫中的焦虑样状态和强迫游泳时的抑郁样情绪,同时,这2个剂量的BPA降低小鼠的社会玩耍和社会交互能力,提示青春期BPA暴露导致的动物社会行为减弱可能与其加重的焦虑抑郁样情绪有关.
作为一种高级动机行为,社会行为具有奖赏价值,动物在与同伴、后代或性伴侣的社会互作中得到奖赏[25].因此,社会动机和奖赏是社会行为的动力.研究发现,与高级动机行为相关的中脑腹侧背盖区-纹状体的DA奖赏通路对社会行为发育至关重要[11].大鼠中脑神经元DA合成限速酶、纹状体DA受体密度均在青春期达到高峰后回落到成年水平,提示DA奖赏系统在青春期社会行为发育中的重要作用[4,11].Trezza等[25]发现社会玩耍伴随着前脑和纹状体DA水平升高,DA传递阻断可减少社会玩耍,而Manduca等[26]发现注射DA再摄取抑制剂安非他命增加社会玩耍行为.笔者的实验室发现,0.40和4.00 mg·kg-1·d-1的BPA暴露上调纹状体DAT表达水平.DAT是一种位于多巴胺能神经元突触前膜上的多巴胺转运蛋白,可将释放至突触间隙的多巴胺主动转运再摄取回突触前.BPA暴露导致DAT蛋白表达水平升高,加快对突触间隙DA的移除而导致纹状体突触前DA能神经纤维末梢DA传递效应减弱.同时,实验发现BPA还上调纹状体DA受体DR1表达水平.脑内DR1被认为在社会行为中起重要作用,抑制纹状体DR1可促进社会互作和社会探究等亲社会行为[27].最近有报道表明,出生前后BPA暴露改变小鼠中脑DA系统和吗啡奖赏活动[4],上调DA转运体基因,上调边缘前脑DR1而下调DR2[28],这与笔者的发现相一致.因此,青春期BPA暴露引起的纹状体DA递质系统功能改变可能是其影响雌性动物社会行为异常的一个原因.
内源性性激素对相关脑区的调节是青春期脑和行为发育性别差异的主要原因.给发育中的脑注射睾酮可使青春期雌鼠的玩耍行为雄性化,脑内ER敲除的小鼠社会探究、攻击和性行为均减少[11].实验发现,低剂量BPA暴露没有明显改变脑和血清的雌二醇水平,但是下调了与社会行为密切相关的纹状体脑区ERβ蛋白表达.雌激素调节脑和行为主要通过ER介导,其中脑功能和非生殖相关行为主要由ERβ介导.在以往的研究中实验室曾发现,长期暴露BPA下调雌鼠ERβ水平[29].因此,青春期BPA暴露可能通过下调脑内ERβ水平而抑制雌激素对社会行为的调节.
4 结 论
综上分析,青春期暴露于低剂量BPA可减少雌鼠社会玩耍,降低其社会探究意愿和社会互作能力.该作用可能与纹状体脑区ERβ水平下调而干扰雌激素对相关脑区DA递质系统活动的调节有关,最后导致社会行为改变.鉴于杏仁核和前额叶与焦虑抑郁情绪相关,今后将进一步探讨其在BPA影响情绪中的作用.本研究发现BPA影响社会行为和情绪的有效剂量为0.40,4.00 mg·kg-1·d-1,根据体表面积换算成人的药理剂量为0.032 4,0.324 0 mg·kg-1·d-1,该剂量远低于现有BPA的无不良效应极限值(50 mg·kg-1·d-1).因此,BPA对神经行为发育的低剂量毒性作用应引起重视.
猜你喜欢
中国体育科技(2022年1期)2022-03-10
中国康复(2021年6期)2021-11-30
青岛大学学报(医学版)(2021年5期)2021-11-17
今日农业(2020年19期)2020-12-14
新世纪智能(英语备考)(2020年6期)2020-08-14
新世纪智能(英语备考)(2020年5期)2020-08-11
做人与处世(2020年10期)2020-06-29
小学生学习指导(低年级)(2019年9期)2019-09-25
科技创新与品牌(2018年5期)2018-07-24
新农村(2018年35期)2018-04-02
