
2017—2018 年广东省H9 亚型禽流感病毒HA 基因序列分析
2019-11-04 03:32吴立炀王福广查云峰许秀琼孙彦伟
中国动物检疫 2019年11期
吴立炀,王福广,查云峰,叶 健,刘 劼,许秀琼,孙彦伟
(广东省动物疫病预防控制中心,广东广州 510230)
H9 亚型禽流感病毒自1966 年首次从北美火鸡中分离到以来,逐渐在世界范围内流行。我国自1994 年在广东省鸡群中首次分离到H9N2 亚型禽流感病毒[1]以来,共发生两次H9N2流感大流行:1998 年首次流行,以Beijing/94 亚群为主;2008年第2 次流行,以Y280 亚群为主[2]。目前,我国H9 亚型禽流感病毒核酸检出率居高不下,以隐性感染为主。此外,H9N2 亚型禽流感病毒与其他亚型流感病毒感染同一宿主并在宿主体内复制过程中,能为其他亚型流感病毒提供内部基因,使其重组为新型流感病毒,如1997 年感染人的H5N1亚型禽流感病毒和2013 年感染人的H7N9 亚型流感病毒[2],这对病毒变异和跨种传播起到了重要作用。
本研究对广东省2017—2018 年检出的13 株H9 亚型禽流感病毒HA 基因进行序列测定以及对基因内部位点进行分析,了解其变异情况,为H9亚型禽流感的有效防控提供参考。
1 材料与方法
1.1 材料
1.1.1 样品 13 份咽/肛拭子样品,为本中心2017—2018 年常规监测项目中检出的部分阳性样品,来源于广东省9 个市的活禽市场。
1.1.2 试剂 核酸抽提试剂盒(磁珠法),购自上海拜诺生物科技有限公司;PrimeScript™ One Step RT-PCR Kit Ver.2、载体PMD-18T 和DH5 α 感受态细胞,均购自宝日医生物技术(北京)有限公司(TAKARA);琼脂糖,购自BIOWEST;切胶回收试剂盒,购自Promega;GenGreen nucleic acid gel stain 显色剂,购自广州盈懋分析测试技术服务有限公司。
1.1.3 HA 基因扩增引物 根据禽流感病毒RT-PCR 检测方法(NY/772—2013),合成1 对甲型流感通用引物:上游引物5'-GGGAGCAAAAGCAGGGG-3';下游引物5'-GGAGTAGAAACAAGGGTGTTTT-3'。引物由上海立菲生物技术有限公司合成。
1.2 方法
1.2.1 病毒RNA 提取 按核酸抽提试剂盒(磁珠法)操作说明书提取RNA。
1.2.2 HA 基因扩增 上下游引物各1 μL(浓度为0.025 nmol/μL),Prime Script 1 step Enzyme Mix 2 μL,2×1 step Buffer 25 μL,RNase Free dH2O 11 μL,样品 RNA 10 μL。扩增条件:50 ℃30 min,1 个循环;94 ℃ 2 min,1 个循环;94 ℃30 s,50 ℃ 30 s,72 ℃ 80 s,34 个循环;72 ℃8 min,1 个循环;4 ℃终止。
1.2.3 PCR 产物回收和转化 对PCR 产物进行凝胶电泳后,在蓝光切胶仪中切下目的片段,按切胶回收试剂盒使用说明书进行目标片段回收;与载体PMD-18T 进行连接,并转化至DH5α 感受态细胞中,通过氨苄培养基筛选得到转化成功菌液,再利用RT-PCR 检测目的片段是否与载体PMD-18T 连接成功。
1.2.4 目的基因核苷酸序列测定和分析 将阳性样品送上海立菲生物技术有限公司测序。使用DNAstar 软件对测序结果进行序列拼接,以在GeneBank 下载的8 株H9 亚型禽流感病毒HA 基因序列作为参考,使用Megalign 软件进行HA 基因序列比对,使用Mega 7.0 软件绘制HA 基因进化树。
2 结果与分析
2.1 HA 基因遗传进化分析
根据Jiang 等[3]建立的遗传进化树分类体系,发现本研究的13 株H9 亚型禽流感病毒均属于h9.4.2.5 分支,同源性介于94.4%~99.6%。而目前的疫苗毒株 A/Chicken/Guangdong/SS/94、A/Chicken/Shanghai/F/98、A/Chicken/Shandong/6/96均属于h9.4.2.2 分支,流行毒株与疫苗毒株同源性为88.2%~90.0%。详见图1。
2.2 潜在糖基化位点分析
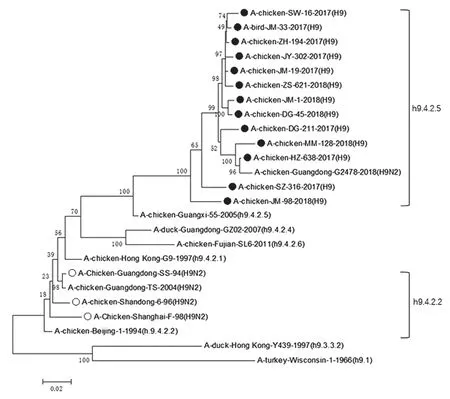
图1 H9 亚型禽流感HA 基因系统进化树
氨基酸序列分析表明,H9 亚型禽流感病毒HA 的潜在糖基化位点一般分别位于29~31、82~84、141~143、218~220、298~300、305~307、313~315、492~494、551~553 aa 等9 个位点上,1~7 位于HA1 上,8~9 位于HA2 上。但与参考毒株A/Chicken/Beijing/1/1994 相比,本研究的13 个毒株均存在218~220 aa 位点的缺失和313~315 aa位点的增加,与参考毒株A/Chicken/Guangdong/G2478/2018 相同。详见表1。
2.3 裂解位点与受体结合位点分析
裂解位点分析发现,13 个毒株裂解位点均为RSSR/GLF,呈低致病性禽流感分子特征。构成受体结合位点(RBS)的7 个氨基酸(109、161、163、191、198、202、203 aa),均 为YWTNTLY。与A/Chicken/Beijing/1/94 相比,均出现V198T 突变,与A/Chicken/Guangxi/55/2005 和A/Chicken/Guangdong/G2478/2018 相同。受体结合位点右缘146~150 位有11 个毒株出现K149N 和A150T 突变,与中国动物卫生与流行病学中心分离到的毒株A/Chicken/Guangdong/G2478/2018 相同,另有2 个毒株出现K149T 突变。受体结合位点左缘232~237 位,13 个毒株均出现Q234L 和Q235M突变。详见表2。
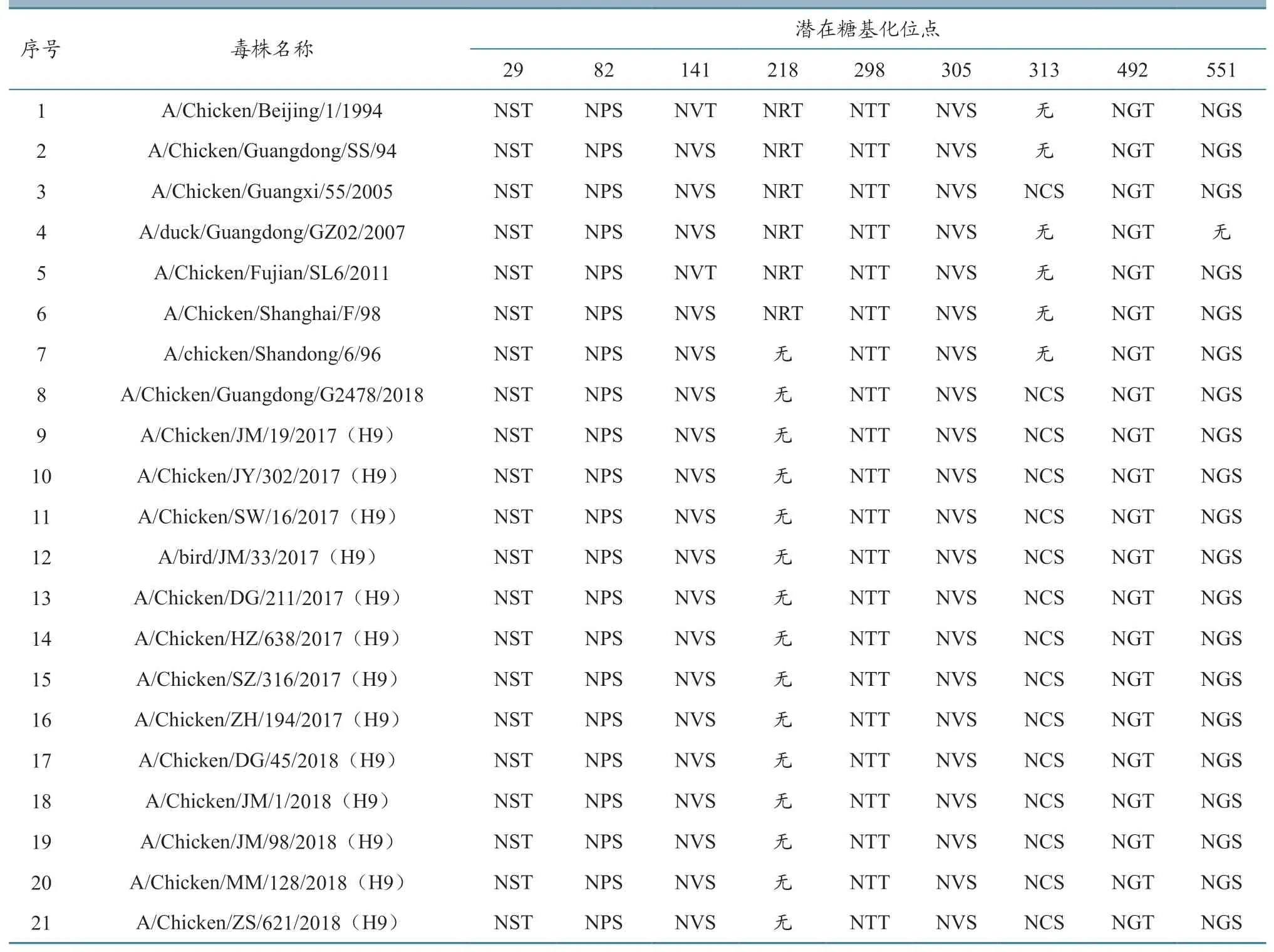
表1 潜在糖基化位点分析结果
2.4 HA 抗原相关位点分析
HA 蛋 白G90E、S145D、D153G、N167G、A168D、T200R、N201D 等氨基酸突变(H9 编码)[4-6]与H9N2 亚型禽流感病毒抗原变异相关。本研究对13 株H9 亚新禽流感病毒的7 个抗原位点进行分析,发现所有毒株均在145、153、167、200 aa 位点出现上述突变,90 aa 位点有12 个毒株出现G90E 突变,168 aa 位点有12 个毒株出现A168N 突变,表明当前H9 亚型禽流感的抗原性发生了较大程度的变异。详见表3。
3 讨论
3.1 当前流行株分类学地位
本研究的13 个毒株均属于h9.4.2.5 分支,表明当前广东省的H9 亚型禽流感优势流行株为h9.4.2.5 分支的毒株。3 个经典疫苗毒株 A/Chicken/Guangdong/SS/94、A/Chicken/Shanghai/F/98、A/Chicken/Shandong/6/96 均属于h9.4.2.2 分支,与流行毒株同源性仅为88.2%~90.0%,表明当前广东省流行毒株与疫苗株出现了较大的遗传差异。
3.2 流行毒株致病性变化的分子基础分析
13 株H9 亚型禽流感病毒的HA 基因序列裂解位点均为RSSR/GLF,呈低致病性禽流感分子特征。糖基化位点主要表现为218~220 aa 位点的缺失和313~315aa 位点的增加。这些糖基化位点的变化是否会影响病毒特性仍有待进一步研究。受体结合位点主要表现为K149N、A150T、V198T、Q234L 和Q235M 位点的突变,与刘彦云等[7]的研究结果相比,RBS 增加了K149N、A150T、Q235M 三个位点的突变。Ha 等[8]研究表明,HA蛋白RBS 的234~236 aa 位点突变为LMG,与人源受体相同,具有与SA-2,6-Gal 结合的能力。本研究中13 个毒株HA 蛋白RBS 的234~236 aa 位点均突变为LMG,具备感染人类的可能性。抗原性相关位点除了201 aa 位点较为保守外,其余6 个位点主要表现为G90E、S145D、D153G、N167G、A168N、T200R 位点的突变,还有168 aa 由天冬氨酸(D)突变为天冬酰胺(N),并成为主要氨基酸。它可能导致H9 亚型禽流感病毒发生抗原漂移以逃避疫苗免疫[9]。2017—2018 年,本中心监测的H9 亚型禽流感抗体合格率分别为80.84%和81.34%,禽群抗体合格率保持在70%以上的有效保护[10]水平,但流通环节H9 亚型禽流感病毒检出率逐年上升,推测目前使用的疫苗与流行毒株匹配度较低,只能提供有限的保护,应考虑以当前的流行毒株研发新的疫苗毒株。

表2 裂解位点与受体结合位点分析结果

表3 HA 抗原相关位点分析结果
猜你喜欢
动物医学进展(2022年9期)2022-11-26
中国生物制品学杂志(2022年3期)2022-05-13
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
复旦学报(医学版)(2021年5期)2021-10-13
皮肤病与性病(2021年3期)2021-07-30
癌症进展(2018年11期)2018-12-30
医学研究杂志(2015年7期)2015-06-22
医学研究杂志(2015年12期)2015-06-10
