
河北地区入侵植物黄顶菊与本地种苍耳光合特性比较
2019-10-25 04:18皇甫超河安徽大学资源与环境工程学院安徽合肥3003农业部环境保护科研监测所天津3009
生物安全学报 2019年3期
贾 璇, 皇甫超河,∗安徽大学资源与环境工程学院,安徽 合肥3003; 农业部环境保护科研监测所,天津3009
生物入侵是目前威胁全球生态环境和经济发展的全球性问题之一。 入侵植物自身的生理特性对于其种群的入侵、生存和扩展至关重要(张晓亚等,2015)。 入侵植物相对于本地植物通常具有更高的叶片N 含量(Nmass),比叶面积(specific leaf area,SLA)和光饱和速率(Pnmax)越高,Nmass就越高,有助于提高入侵植物生物量,从而获得相对于本地植物更高的竞争优势。 Feng et al. (2007)的“N 分配的进化假说”认为入侵植物的入侵机制与植物的光合作用有很大关系,相对于原产地种群,入侵到中国的入侵种群有着较高的光合氮利用效率(photosynthetic nitrogen use efficiency,PNUE)、光合能量利用效率(photosynthetic energy use efficiency,PEUE)以及更高的能量利用返还策略(a quicker return energy-use strategy)。 另外,入侵植物增加了对光合机制的N 分配,减少了对细胞壁等防御结构的N 分配,导致光饱和速率的增加,可能也增加了PEUE等(Liu et al.,2017)。 在我国现有的入侵植物中菊科植物所占的种数最多,我国已经确定的230 种入侵植物中有59 种隶属于菊科(罗雪晶等,2017)。由于具有相同的进化历史和生长策略,开展同科属近缘入侵种和土著种比较研究有助于解释入侵物种快速扩散的生理生态学机制(John & John,2011)。
黄顶菊Flaveria bidentis (L.) Kuntze 属菊科黄顶菊属,一年生草本植物,原产于南美洲,分布在西印度群岛、墨西哥和美国的南部,后来又传播到欧洲的英国、法国,澳大利亚,非洲的埃及和南非以及亚洲的日本等地(刘全儒,2005)。 黄顶菊于2001年在我国天津南开和河北衡水市首次被发现,传入年份及传入途径不清楚(皇甫超河等,2009)。 黄顶菊入侵我国后,凭借其旺盛的繁殖能力和广泛的适应性迅速得到传播,并通过与本地物种的竞争或占据本地物种的生态位来排挤本地植物,降低入侵地物种多样性水平和生态系统的功能(刘宁,2013)。目前,小飞蓬Conyza canadensis (L.) Cronq(王晓红和纪明山,2013)、少花蒺藜草Cenchrus pauciflorus Benth(王坤芳等,2016)、马缨丹Lantana camara L.(朱慧和马瑞君,2009)与其伴生物种的光合特性均有报道,研究认为,入侵种有较高的光合速率和物质积累能力以及较高的生产力。 皇甫超河等(2009)初步研究了黄顶菊及其4 种伴生杂草的光合特性。 前人的研究均在人工种植条件下开展,缺乏在自然生境下入侵种与本土近缘物种光合特性的报道,且关于黄顶菊对外部光强环境响应模式尚无研究。 基于此,本文推测黄顶菊能够在河北地区成功入侵与其光合特性有重要关系,黄顶菊相对本地种具有较高的气体交换参数、光合能量利用效率等,从而具有对环境资源的利用效率更高、捕获能力与利用能力更高等特点。 本研究选取河北邯郸、衡水和沧州地区黄顶菊的典型重发生区,以共生的菊科本地植物苍耳Xanthium sibiricum Patrin ex Widder 为对照,测定二者相关光合参数,比较分析二者的光合特性及叶片特性之间几种重要指标的差异。 苍耳亦为菊科一年生草本植物,在黄顶菊入侵区是具有代表性的土著共生种(屠臣阳等,2013)。 因此,探讨二者对光强变化的响应模式和揭示黄顶菊环境适应性的生理生态学机制具有重要参考价值。
1 材料与方法
1.1 材料
分别于2014 年9 月7、8、9 日在河北3 个地区进行数据测定和样品采集。 所选生境均为黄顶菊和苍耳混生,其他伴生植物包括灰绿黎Chenopodium glaucum L.、反枝苋Amaranthus retroflexus L.、狗尾草Setaria viridis (L.) Beauv.等。
在河北省邯郸市永年县临洺关镇洺河滩(36°47′51″N, 114°28′48″E,海拔60 m)进行邯郸地区的植物数据测定和采样,此地为黄顶菊重发生区。 此区域有比较典型的暖温带半湿润大陆性季风气候,日照充足,四季分明,春季干旱少雨,夏季炎热多雨,秋季温和凉爽,冬季寒冷干燥。 年平均温度13.5 ℃,全年无霜期200 d,年均日照时数2557 h。 土壤理化性质为:土壤pH 值7.61,有机质14.66 g·kg-1,全N 1.63 g·kg-1,全P 0.70 g·kg-1,土壤类型为盐渍土。
在河北省衡水市衡水湖畔(37°33′29″N, 115°34′49″E, 海拔20 m)进行衡水地区的数据测定和植物采样,此地区为黄顶菊重发生区。 此区域冷暖干湿差异较大,属于暖温半干旱型大陆性季风气候。 年平均温度13.0 ℃,年平均降水量518.9 mm,年平均日照时数2197 h,四季分明。 土壤理化性质为:土壤pH 值8.30,有机质6.92 g·kg-1,全N 0.78 g·kg-1,全P 0.60 g·kg-1,土壤类型为潮土。
在河北省沧州市献县陌南村边(38°15′2″N,115°58′17″E,海拔10 m)进行沧州地区的数据测定和植物采样,同样为黄顶菊重发生区。 该区气候特点是四季分明,光照充足,属于暖温带大陆季风气候,年平均温度12.5 ℃,年平均降水581 mm。 年积温4785 ℃,无霜期181 d。 土壤理化性质为:土壤pH 值8.31,有机质10.42 g·kg-1,全N 1.10 g·kg-1,全P 0.58 g·kg-1,土壤类型为盐渍土。
1.2 方法
在邯郸、衡水、沧州3 个试验区域都以苍耳和黄顶菊分别随机设置一个面积为10 m×10 m 的试验区组,同一采样时间,在各地区设置的区组内设置5 个1 m×1 m 的小样方,此5 个样方必须均匀分布在每个区组内部;同时在这5 个小样方内随机抽取10 株生长较好并且大小、长势均高度一致的具有代表性的植株,在此10 株植株上分别采集30 片健康完整且大小、颜色相一致的成熟叶片。 2 种植物均采用一致的取样方法,并且在各地区各试验区组内的5 个小样方中2 种样品的采集都设置5 次的重复采样。 采样的同时将采样植物根围0 ~10 cm 的土壤用土壤采样器进行采集,保存好带回实验室之后进行土壤的物理化学性质分析。 采用Li-3100A 叶面积仪(Li-Cor, Lincoln, Nebraska, USA)测定样品的叶片面积,然后将叶片放在冰盒中进行低温保存,用于测定叶片中的叶绿素含量以及叶片N、P 含量等理化指标。
光合—光强(Pn-PAR)响应测定:在样方中选取处于营养生长期生长状况良好的植株,选取完全展开、生长成熟的叶片,用Li-6400 便携式光合测定仪(Li-Cor Inc., Lincoln, NE, USA)测定叶片的净光合速率(net photosynthetic rate, Pn)、蒸腾速率(transpiration rate, Tr)、气孔导度(stomatal conductance, Gs) 等指标(吴彦琼和胡玉佳,2004)。 由于河北3 个地区的年光辐射量在1630 ~1860 kw·h·m-1范围内,因此本研究将有效光合辐射梯度分别设置为2000、1500、1000、800、500、300、100、50 和0 μmol·m-1·s-1,利用自动测量程序分别进行光合—光强响应的测定(朱慧和马瑞君,2009)。 测定时室温均设置为25 ℃,CO2浓度控制在400 μmol·mol-1,测定时保持叶片自然生长角度不变,重复3 次,每区组叶片测定重复5 次,最终结果以测定结果的平均值为准。
叶绿素含量的测定:用去离子水清洗采取的叶片,清洗过后的叶片再用打孔器取片之后切成长细丝状,用80%的丙酮浸叶片至其完全变为白色后取出,分别在波长为663、645 nm 处读取光密度(记为D663nm、D645nm)。
叶片元素含量的测定:植物样品于105 ℃下杀青30 min,然后在65 ℃下烘干直到其质量不再发生变化,将烘干后的植物叶片粉碎过筛,混合均匀后保存备用。 分别用开氏消煮法和钼锑抗比色法测定植物叶片的全N 和全P(顾大形等,2011; 鲁如坤,2000; Sparks et al.,1996)。
叶片灰分含量的测定:将叶片放在马弗炉中,在500 ℃下灼烧6 h 后,称量剩余残渣重量并记录。干重热值的测定采用氧弹式热量计(HWR-15E,上海上立检测仪器厂):取0.5 g 植物样品粉末,将样品完全燃烧同时测定热值,5 个重复,以其平均值作为最终测定的干重热值(Williams et al.,1987)。
将带回实验室的土壤样品自然风干后,去除土壤之外的杂物后进行研磨,研磨充分后分别过孔径为0.149 和0.841 mm 的孔筛,将过筛后保留的土壤采用水合热重铬酸钾氧化—比色法进行有机碳测定;土壤中的和采用流动注射分析仪(QC8000, Lachat, USA)测定;土壤中全N 和全P 的测定与上述叶片全N 和全P 的测定方法相同。
1.3 数据处理
参考魏子上等(2017)的数据处理方法,光合—光强响应曲线采用直角双曲线修正模型进行拟合(叶子飘和于强,2008; Ye,2007):Pn=αI(1-βI)/(1+γI)-Rd。 式中:Pn为净光合速率(μmol·m-2·s-1) ;I 为光合有效辐射(μmol·m-2·s-1);α 为表观量子效率;β 和γ 为系数; Rd为暗呼吸速率(mg·h-1·g-1)。
水分利用率(water use efficiency,WUE)(高丽等,2009):Pr=Pn+Tr,WUE=Pr/Tr。 式中:Pr为光合速率(μmol·m-2·s-1);Tr为蒸腾速率(mmol·m-2·s-1)。
叶绿素含量(舒展等,2010):ρ(Chla)= 12.7D665nm- 2. 69D645nm,ρ(Chlb) = 22. 9D645nm- 4.68 D663nm,ρ(Chl)=ρ(Chla)+ρ(Chlb)。 式中:ρ(Chla)为叶绿素a 的含量(mg·L-1);ρ(Chlb)为叶绿素b的含量(mg·L-1);ρ(Chl)为叶绿素总含量(mg·L-1)。
叶片灰分含量:Hc= CV/(1-Ash),CCmass=[(0.06968 Hc- 0.065) × 1 -Ash) + 7.5(k ×ON/14.0067]/EG(屠 臣 阳 等,2013; Williams et al.,1987)。 式中:Hc为去灰分热值(kJ·g-1);Ash为灰分含量(%);CV 为干重热值(kJ·g-1);CCmass为叶片单位质量建成成本(gglucose·g-1);ON 为有机N含量(%);EG为生长效率。 不同植物的平均叶片生长效率为0.87(Penning et al., 1974)。
叶片比叶面积:SLA = ULA/DW,Harea= Nmass/SLA,CCarea=CCmass/SLA,PEUE=Pnmax/CCarea(Funk& Vitousek,2007),PNUE=Pnmax/Narea(Field & Mooney,1986)。 式中:SLA 为叶片比叶面积(cm2·g-1);ULA 为单位叶片面积(cm2);DW 为叶片干质量(g);CCarea为叶片单位面积建成成本(gglucose·m-2);Nmass为叶片单位质量N 含量(mg·g-1);Narea为叶片单位面积N 含量(g·m-2);PEUE 为光合能量利用效率(μmol·gglucose-1·s-1);PNUE 为光合氮利用效率(μmol·g-1·s-1)。
以上所有数据采用SPSS l6.0 和Excel 2007 进行统计分析,用Origin 8.0 进行图形制作。 用Pearson 相关系数分析法进行相关性分析,并进行双尾显著性检验。 叶片特性指标的比较中得到的数据为配对t 检验的结果。 采用单因素方差分析法进行2 个物种间均值的方差分析,并且采用最小显著差数法对实验结果进行差异显著性比较(P=0.05)。
2 结果与分析
2.1 河北3 个地区黄顶菊与苍耳对有效光合辐射的响应
2.1.1 邯郸地区黄顶菊与苍耳对有效光合辐射的响应 邯郸地区的2 种植物随着有效光合辐射的增加,其Pn、Gs、Tr和WUE 均呈增加趋势,且除Tr之外,黄顶菊气体交换参数的最大值显然高于苍耳,而苍耳的Tr曲线上升趋势要高于黄顶菊;但在有效光合辐射大于1000 μmol·m-2·s-1时,苍耳的Gs、Tr和WUE 趋于一个稳定值,并没有继续随着有效光合辐射的增强而增加;而黄顶菊的Pn、Tr和WUE 在有效光合辐射达到1000 μmol·m-2·s-1时依然有持续增加的现象(图1)。
2.1.2 衡水地区黄顶菊与苍耳对有效光合辐射的响应 衡水地区的2 种菊科植物随着有效光合辐射的增加,其Pn、Gs、Tr和WUE 均表现为增加的趋势,同时黄顶菊的各项指标在同样的有效光合辐射下均明显大于苍耳,且Pn差异最为显著;当有效光合辐射到达约1000 μmol·m-2·s-1时,除Tr之外,随着有效光辐射的继续增强,苍耳的Pn、Gs和WUE也呈现了与邯郸地区相同的趋势,即趋于一个稳定值(图2)。
2.1.3 沧州地区黄顶菊与苍耳对有效光合辐射的响应 沧州地区的2 种植物随着有效光合辐射的增加,其Pn、Gs、Tr和WUE 也均呈增加趋势,且苍耳的Pn、Gs、Tr和WUE 曲线趋势和衡水地区相同;两者的Gs差异显著,黄顶菊有着较高的Gs,Tr曲线趋势却近似一条直线,与前2 个地区不大相同,说明此时黄顶菊的蒸腾速率较高,推测可能是因为测定时外界温度过高引起的(图3)。
如图所示,3 个地区黄顶菊和苍耳的Pn、Gs、Tr和WUE 虽然都随着有效光辐射的增加而增高,但黄顶菊的Pn、Gs、Tr和WUE 在同一有效光辐射的条件下始终大于苍耳。 除邯郸地区的Tr-PAR 曲线之外,河北3 个地区黄顶菊的气体交换参数Pn、Gs、Tr和WUE 随有效光合辐射的增加幅度要高于苍耳。 由此可见,黄顶菊的气体交换参数高于苍耳。
2.1.4 3 个地区2 种植物的光合响应参数 采用直角双曲线修正模型计算出河北3 个地区中黄顶菊和苍耳的光合响应参数(表1)。 邯郸和衡水地区的黄顶菊光饱和点(light saturation point,LSP)均显著高于苍耳;这2 种菊科植物的LSP 均大于1200 μmol·m-2·s-1。 3 个地区黄顶菊的光补偿点(light compensation point,LCP)均显著低于苍耳,除衡水地区外,其他2 个地区的黄顶菊的表观量子效率(apparent quantum yield,AQY)也均显著低于苍耳;3 个地区黄顶菊的Pnmax均显著高于苍耳,分别高出各自本地种的42.8%、221.8%和122.9%。

图1 邯郸地区2 种植物光合—光响应曲线的比较Fig.1 Comparison of Pn-PAR curve of F. bidentis and X. sibiricum in Handan
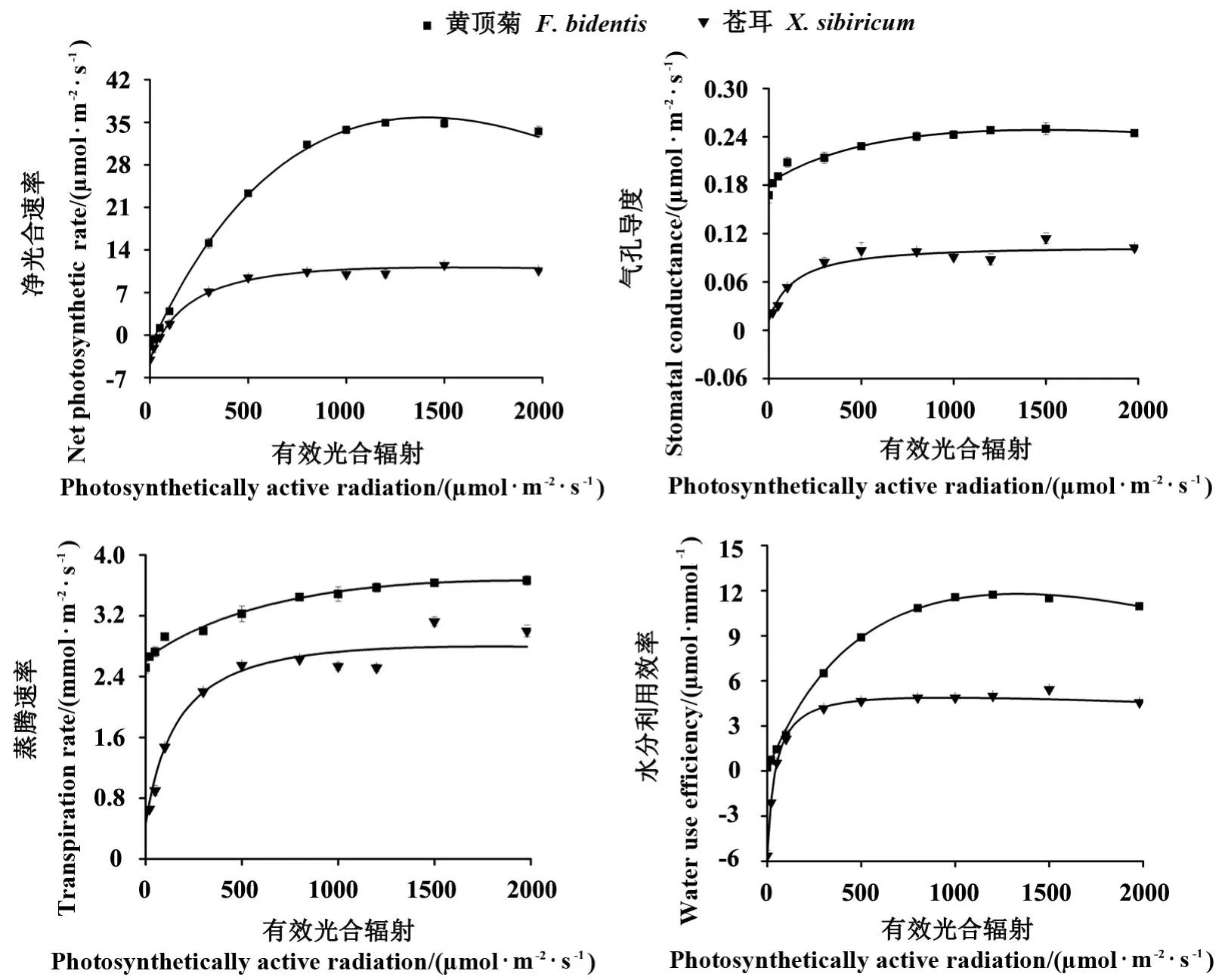
图2 衡水地区2 种植物光合—光响应曲线的比较Fig.2 Comparison of Pn-PAR curve of F. bidentis and X. sibiricum in Hengshui
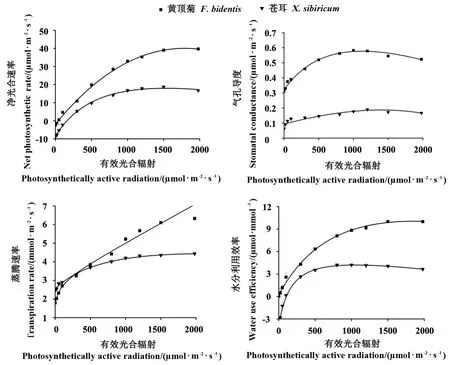
图3 沧州地区2 种植物光合—光响应曲线的比较Fig.3 Comparison of Pn-PAR curve of F. bidentis and X. sibiricum in Cangzhou

表1 河北3 个地区黄顶菊与苍耳的光合响应特征参数Table 1 Photosynthetic parameters of F. bidentis and X. sibiricum in three areas in Hebei Province
2.2 黄顶菊与苍耳叶片特性指标的比较
对3 个地区的黄顶菊及其苍耳的叶片特性指标进行测定(表2),3 个地区的入侵植物黄顶菊和苍耳叶绿素含量差异不显著;除邯郸之外,其他2个地区黄顶菊的SLA、PNUE 均要显著高于苍耳(P<0.05),且3 个地区黄顶菊的Nmass、Pmass、PEUE 均显著高于苍耳(P<0.05)。
2.3 黄顶菊与苍耳最大净光合速率与叶片特性指标的相关性分析
对3 个地区2 种植物的气体交换参数和叶片特性指标进行相关性分析(表3),结果表明,植物Pnmax与叶片CCmass呈显著正相关(P <0.05),与Nmass、Pmass、PEUE、PNUE 呈 极 显 著 正 相 关(P <0.01);Nmass与SLA、CCmass、Pmass呈极显著正相关(P<0.01);SLA 与PEUE 呈显著正相关(P<0.05),与PNUE 呈极显著正相关(P<0.01)。
3 讨论
分析河北3 个地区的黄顶菊和苍耳对有效光辐射的响应,发现3 个地区的黄顶菊的净光合速率、气孔导度、蒸腾速率和水分利用率均大于苍耳,气孔不但是CO2进出的通道,也控制O2和水蒸汽的扩散。 所以,植物气孔的开闭不仅与植物的光合作用有关,也影响植物的呼吸作用和蒸腾作用(陈新微等,2016)。 相对于苍耳,黄顶菊较高的气孔导度表明在相同的日照环境条件下,黄顶菊能够固定更多的CO2,具有更高的光合利用效率。 且随着有效光辐射的增加,黄顶菊的净光合速率、蒸腾速率和水分利用率均增长。 而当有效光辐射到达一定数值后,苍耳的净光合速率、蒸腾速率和水分利用率逐渐趋于平缓,未继续上升。 本研究显示,河北3个地区的黄顶菊的LSP 均大于苍耳,且Pnmax也显著高于苍耳。 因此,黄顶菊对于高的光照强度具有更强的适应性,这可能是其能够在河北地区成功入侵的原因之一。

表2 河北3 个地区2 种植物叶片特性的比较Table 2 Comparison of leaf traits of F. bidentis and X. sibiricum in three areas in Hebei Province

表3 2 种植物Pnmax与叶片特性指标之间的相关性Table 3 Correlation coefficient among Pnmax and leaf characteristics of F. bidentis and X. sibiricum
N 可以提高植物对C 的同化效率,提高其光合作用能力。 高N 能够增加植物光合过程中的最大羧化和电子传递速率、增大Pnmax,从而提高对光合有效辐射的利用和适应能力。 N 也是构成蛋白质的主要成分,对茎叶生长、果实发育起重要作用,尤其在结果期,植物对N 的吸收达到高峰。 因此,高N 能够为自身提供竞争优势。 本研究中,除邯郸地区之外,其他2 个地区黄顶菊的PNUE 显著高于苍耳,3 个地区黄顶菊的Nmass显著高于本地种,表明黄顶菊对于N 的利用能力高于苍耳,具有较高的光合N 利用效率。 CCmass是衡量植物叶片建成所需能量的重要指标,入侵植物较低的CCmass可能会增加其竞争优势(屠臣阳等,2013)。 本研究中,河北3个地区的黄顶菊的CCmass均低于苍耳,体现了黄顶菊更高的能量利用策略。 而SLA 较高的植物能将更多的N 分配到光合作用中,进而影响植株的相对生长速率、CCmass和Pnmass等(Callaway et al.,2004)。研究表明,黄顶菊能够凭借较高的SLA 和较低的CCmass,将N 更多地分配到光合作用中去,从而提高其光合N 利用效率,这也是黄顶菊能够成功入侵的重要原因。
综合以上分析,在河北地区,相比本地种苍耳,黄顶菊均有着较高的光合特性和能量利用指标,在同一环境条件下,黄顶菊有着更高的光合速率、PEUE 以及PUNE,及更高的光捕获能力和利用能力,对高的光照强度具有强的适应性,因而能迅速在河北地区侵占本地物种资源与生境成功入侵。研究表明,土壤中N 的增加能够提高入侵植物三叶鬼针草Bidens pilosa L.的相对生长速率(潘玉梅等,2012)。 将来需要进一步开展N 肥添加实验,探讨黄顶菊高的光合N 利用效率与土壤中的N 水平的关系。
猜你喜欢
肉类研究(2022年7期)2022-08-05
河北果树(2021年4期)2021-12-02
现代畜牧科技(2021年4期)2021-07-21
大众文艺(2020年20期)2020-11-05
幼儿智力世界(2020年10期)2020-10-28
流行色(2020年9期)2020-07-16
作文大王·低年级(2020年2期)2020-03-13
学生天地(2019年30期)2019-08-25
家庭影院技术(2018年9期)2018-11-02
CHIP新电脑(2017年6期)2017-06-19
