
基因水平转移在根瘤菌进化中的研究进展
2019-10-24 09:11:08陈雪莲江高飞钟增涛
生物技术通报 2019年10期
陈雪莲 江高飞 钟增涛
(南京农业大学生命科学学院,南京 210095)
根瘤菌是一类革兰氏阴性土壤细菌,能够与大多数豆科植物共生固氮。根瘤菌与植物共生形成根瘤或茎瘤,在根瘤内分化成类菌体后能够将空气中的氮气通过生物固氮作用转化为铵。植物进一步将铵转化为氨基酸等氮源物质,满足其生长繁殖需要[1]。豆科植物是植物界最大的一个科,约有650属19 700种,大多数豆科植物可以与根瘤菌建立共生固氮体系[2]。由于根瘤菌的宿主专一性较强,如何扩大根瘤菌在豆科植株上共生固氮的宿主范围,最终实现在非豆科植物上的共生固氮是根瘤菌研究的最终目标[3],也是生物固氮研究的目的之一。
1 基因水平转移(HGT)
原核生物具有从环境或其他生物体基因组获取部分DNA的独特能力,并在称为基因水平转移(Horizontal gene transfer,HGT)的过程中将其整合到自身基因组中[4]。基因水平转移是相对垂直进化(通过亲代传给子代)而言的,可以让细菌快速获得新的遗传性状从而促进细菌的进化。基因水平转移在微生物进化过程中发挥着重要的作用,它打破了遗传依赖亲缘关系的界限,使遗传物质能够在不同的物种之间进行交换,原核生物通过基因水平转移的方式可提高对环境的适应性,并具备遗传的多样性[5-6]。通过这种方式,微生物可以获得新的基因型和表型,例如毒力、抗生素抗性、代谢和共生等[6]。基因水平转移的主要类型分别是:(1)转化:从环境中吸收外源DNA的能力;(2)转导:通过噬菌体介导将DNA从一个细胞转移到另一个细胞;(3)接合:通过质粒或者接合转移元件将DNA从供体细胞向受体细胞进行接触型转移。转导和接合都是由遗传元件介导的,但是转导不需要细菌间的接触。细胞内或细胞间的可移动遗传原件(Mobile genetic element,MGE)主要有:转座子类原件、噬菌体/病毒、接合型质粒和整合与接合原件(Integrative and Conjugative Elements,ICEs)。ICEs含有编码接合相关的基因以及复杂的调控系统以调控其从染色体上切离和接合转移[3],它们能整合在染色体上并随着染色体的复制而复制。在特定条件诱导下,ICEs从染色体上切离并环化,复制后通过接合的方式转移到新的宿主细胞中,供受体细胞中环化的ICEs均可再次整合到染色体上。与温和噬菌体类似,ICE通常整合在宿主染色体内的一个或多个特定位点(通常是氨酰tRNA),通过复制和解离使ICE向其它细胞转移[7]。切离的ICE通常呈环状,通过接合的方式向受体细胞发生转移[8]。ICEs不同于质粒介导的接合转移,它不能以染色体外的状态独立存在,且不能独立复制[9],因此重新整合到宿主染色体对于其遗传稳定性至关重要。相对于质粒介导的基因水平转移,ICEs因为能整合到染色体上,因此遗传稳定性更高,能更好地促进受体细胞基因组的遗传进化;同时其所含基因簇在功能上具有较强的一致性且种类多样,因此具有更加丰富的遗传多样性。
2 根瘤菌中的基因水平转移方式
根瘤菌的共生固氮相关基因是以基因簇的形式存在的,主要存在于共生质粒或者是染色体的共生岛上。在根瘤菌属(Rhizobium)和中华根瘤菌属(Sinorhizobium)中主要是以共生质粒的形式存在;在大豆根瘤菌属(Bradyrhizobium)和固氮根瘤菌属(Azorhizobium)中主要以共生岛的形式存在;在中慢生根瘤菌属(Mesorhizobium)中,这两种形式都存在。无论是共生岛还是共生质粒均能发生水平转移。
2.1 共生质粒
根瘤菌属和中华根瘤菌属的根瘤菌中参与共生固氮的结瘤基因和固氮基因,位于可转移的共生大质粒上(pSym),如苜蓿中华根瘤菌Sinorhizbium meliloti1021的共生质粒pSymA等,质粒丢失会导致根瘤菌失去共生能力[10]。菜豆根瘤菌Rhizobium etliCFN42菌株含有6个质粒(p42a-p42f),其大小范围为185-643 kb。其中质粒p42d携带结瘤和固氮所需的大部分信息,而p42a对于pSym的接合转移是必不可少的[11]。当菜豆根瘤菌Rhizobium etliCFN42生长到高细胞浓度时,细菌可以激活p42a 的接合转移,该过程依赖于群体感应系统调控。p42a上的TraI在高细胞浓度时产生的3-oxo-C8-HSL能够与TraR结合,使其形成二聚体激活traI的表达,产生级联放大的作用,从而促进接合转移相关基因的表达并促进pSym向土壤细菌中转移[12]。
2.2 共生岛
基因岛于1990年被Hacker等[13]在Escherichia coli536和E. coliJ96中首次发现,是在基因组上存在着的一段可转移的DNA片段,并将这段两端含重复序列的DNA片段称为毒力DNA岛。基因岛一般整合在基因组特定的氨酰tRNA位点,长度为10-600 kb,该区域的GC含量较低,基因岛上含有整合酶、转座酶。基因岛又根据其携带的功能基因的不同,分为环境适应性岛、抗性岛[14]、代谢岛、毒力岛[15]、耐药性岛和共生岛等。
共生岛首次被鉴定是在百脉根根瘤菌(Mesorhizobium lotiICMP3153)中,大小为500 kb的基因岛,其整合在Phe-tRNA基因的3'末端,内部含有P4整合酶[16],还含有共生固氮相关的nod、nif基因簇[17]。大豆根瘤菌(Bradyrhizobium japonicumUSDA110)全基因组测序结果确定了该菌株含有一个长度为681 kb的共生岛。在其基因组中鉴定了167个转座酶,其中有100个位于共生岛上[18]。M. lotiMAFF303099的共生岛上包含580个基因,其中111个与转座功能相关[19]。M. loti的共生岛ICEMISymR7N是一个502 kb的整合和结合元件(ICE),它可向土壤中的非共生百脉根根瘤菌转移并赋予它们与百脉根(Lotus corniculatusLinn.)共生固氮的能力[20]。有趣的是,该共生岛的转移频率受细菌群体感应系统调控。当细胞浓度高的时候,百脉根根瘤菌的TraR被激活,然后激活自体诱导物合成酶基因traI的表达,使得traI2基因下游的msi172和171发生共转录,通过编程式核糖体移码阅读可分别翻译出Msi172和Msi172-171(FseA),从而激活T4SS(Ⅳ分泌系统),促进共生岛的接合转移。当低细胞密度时,traI2被抑制,因此FseA的表达被抑制,共生岛的接合转移也被抑制[21]。
Azorhizobium caulinodans中含有一个87.6 kb的共生岛(ICEAC),它能够向不同属的根瘤菌中转移,一般整合到受体根瘤菌的Gly-tRNA位点[22]。不同于百脉根根瘤菌的共生岛只能在种内水平上发生转移,田菁根瘤菌(A. caulinodansORS571)的共生岛能分别转移到中慢生华癸根瘤菌(Mesorhizobium huakuii7653R,93)、 百 脉 根 根 瘤 菌(M.lotiNZP2213)、天山根瘤菌(Mesorhizobium. tianshanenseCCBAU3306)及草木樨根瘤菌(Sinorhizobium medicaeUSDA1037)中;受体根瘤菌获得共生岛后,也获得了在毛萼田菁上结根瘤和茎瘤的能力[22]。因为田菁根瘤菌没有AHLs类群体感应系统,所以该共生岛的水平转移不受细胞密度影响,主要受共生岛上整合酶和接合转移相关的酶影响(图1)。
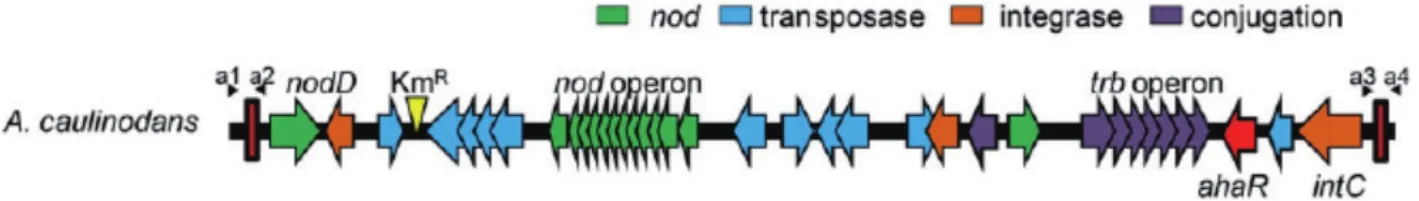
图1 A. caulinodans共生岛的基因结构[22]
3 根瘤菌的进化
目前根瘤菌的分类主要通过分子生物学的方法,包括基因组的GC含量以及16S rRNA测序。目前已经命名的根瘤菌主要分布在α变形菌纲和β变形菌纲的18个属238个种[23];如α变形菌纲中的根瘤菌属(Rhizobium)、伴根瘤菌属(Pararhizobium)、新根瘤菌属(Neorhizobium)、剑菌属(Ensifer)——包含原来的中华根瘤菌属,中慢生根瘤菌属(Mesorhizobium)、固氮根瘤菌属(Azorhizobium)及慢 生 根瘤菌 属(Bradyrhizobium) 等[24];β变形菌纲中的伯克氏菌属(Burkholderia)和贪铜菌属(Cupriavidus)[25]等;但在γ变形菌纲中,目前只发现一种根瘤菌,属于假单胞菌属(Pseudomonas)。随着研究的深入,根瘤菌的所属范围也在逐步扩大。
与许多在特定生态位定殖的细菌不同,根瘤菌在系统发育树中的分布是分散的。这种分散性不仅体现在纲、目等较高的分类水平上,甚至同一个属或种水平上也可以包含根瘤菌和非根瘤菌。通过比较分析α和β变形杆菌纲不同根瘤菌的基因组,除了部分nod/nif基因相对保守外,其它共有的基因数目不足根瘤菌平均编码基因数目的10%,低于系统进化中相似物种核心基因数占比15%的平均值[3]。
在根瘤菌的进化研究中,多以关键的结瘤基因簇(nod)和固氮基因(nif)作为研究对象[26],它们大多存在于共生质粒或共生岛上,可以向其它细菌发生水平转移。在龙舌兰的根际,分离到多种不能结瘤的根瘤菌属(Rhizobium)菌株,它们不含nod基因但是含有nif基因[27]。固氮的起源远早于结瘤,nif基因的分布也更加广泛[28],因此根瘤菌很有可能是这些非共生原始菌株通过基因水平转移获得结瘤基因进化而来[29]。根瘤菌的多点起源假说表明不同分类地位的非根瘤菌可以多次、独立地通过获得共生基因逐步进化为根瘤菌[30]。
3.1 由根瘤菌向非共生根瘤菌转移
目前关于根瘤菌进化的研究多采用比较基因组学和适应性进化的方法。有研究表明,在自然条件下百脉根根瘤菌的共生元件可以向其它非共生根瘤菌中转移,并使其获得在百脉根上的结瘤能力[20]。当不同属的根瘤菌获得田菁根瘤菌的共生岛后,也同时获得了新的能够在田菁上共生固氮的能力[22]。M.lotiR7A的共生岛长502 kb,可向同属非共生的3个种转移,并且使它们能够在百脉根上结瘤[16]。
3.2 由根瘤菌向非根瘤菌转移
从中国台湾相思豆的根瘤中分离出一种根瘤菌D5,其与假单胞菌16S rRNA同源性为99%,其nodA基因与9种慢生根瘤菌属(Bradyrhizobium)具有95%同源性[31],可能是假单胞菌获得根瘤菌的共生岛后进化而来[32]。在野豌豆的根瘤中也分离到了一株克雷伯氏菌属(Klebsiella)的菌株,它能够成功侵染野豌豆并形成具有固氮能力的根瘤[33]。比较基因组学研究发现,根瘤菌Cupriavidus taiwanensisLMG19424和腐生菌Cupriavidus eutrophusH16的基因组具有高度的相似性[3]。Marchetti等[34]通过将C.taiwanensisLMG19424的0.55 Mb共生质粒转移到与其遗传背景相似的青枯菌Ralstonia solanacearumGMI1000中,最终筛选到3个能够在含羞草上结瘤的青枯菌菌株。通过适应性进化的方法筛选出2个对于该菌株从致病型向共生型的转变至关重要的突变株。其中hrcV基因的突变,能够提高突变株的结瘤和早期侵染能力,而毒力蛋白因子hrpG的失活则允许该菌株发生细胞内侵染。虽然在实验室条件下菌株获得了一定的共生性状,但还没有共生固氮表型,因此不能算作真正意义上的根瘤菌[35]。
4 影响根瘤菌基因水平转移的方式
基因水平转移有一定的排它性,亲缘关系越近,这种排他性也就相对较小,基因的交流也就越容易。有研究表明发生基因水平转移的供受体在基因组GC含量上非常相似,大部分差异都小于5%,表明供受体细胞间基因组GC含量的差异对基因转移产生阻碍[36]。此外,供受体基因组的相似性与基因水平转移频率呈正相关[36],这表明基因水平转移在亲缘关系更近的物种中发生的频率更高。转导被认为是距离最长的基因水平转移机制,因为它需要噬菌体介导[37]。Tao等[3]在各种遗传体系(包括微生物染色体、质粒和噬菌体基因组)中重建了共有蛋白家族网络,经过比较发现在自然界中接合比转导发生的频率更高。
异源结瘤基因之间相互抑制可能是根瘤菌基因水平转移的障碍之一。敲除M. huakuii93接合子中ORS571共生岛上的nodD基因后并不影响其在毛萼田菁上结瘤,因为M. huakuii93中的nodD可以弥补共生岛上的缺失[22]。但是缪礼鸿等[38]研究发现费氏中华根瘤菌(Sinorhizobium fredii)HN01SR接受了豌豆根瘤菌(Rhizobium leguminosarum bv viciae)1229的共生质粒后并不能在豌豆上结瘤,只有消除了HN01SR自身的共生质粒,才能够在豌豆上结瘤。张学贤等[39]研究也发现紫云英根瘤菌得到了豌豆根瘤菌共生质粒之后,只有消除本身的共生质粒的才能够在豌豆上结瘤。可见,不同遗传体系下,不同结瘤基因间的相容性对根瘤菌宿主范围的扩大至关重要。
在根瘤菌-豆科植物共生中,类黄酮类化合物通过吸引细菌并诱导NodD的活性从而激活结瘤因子合成基因的表达发挥重要的作用[40-41]。研究表明田菁根瘤菌ORS571在感知到类黄酮类信号分子柚皮素后,LuxRI家族蛋白AhaR与柚皮素结合后形成二聚体,可以特异性结合到整合酶intC的启动子,进而激活intC表达,诱导田菁根瘤菌共生岛ICEAC水平转移[22]。因此,柚皮素可以提高田菁根瘤菌10-20倍的共生岛转移频率。ICEMlSymR7A切除和转移受LuxRI家族自诱导/群体感应(QS)系统的正调节[42]。在获得外源共生固氮遗传物质后,细菌是否能进化成为根瘤菌还需要经过宿主体内长期的选择压力筛选(图2)。将田菁根瘤菌ORS571的共生岛转移到土壤自生固氮菌中,在侵染田菁的初期并不能形成根瘤或茎瘤,通过适应性进化的方法,经过三轮生态循环筛选,获得了可以结瘤并具有一定固氮酶活性的突变菌株(未发表数据),该结果说明土壤细菌获得共生固氮基因后还需要与宿主植物长期进化才能成为根瘤菌。
5 建立共生体系时可能存在的问题
根瘤菌大多数时候在土壤中以腐生的方式存在,只有当遇到合适的宿主植物后,才会进入共生状态,形成根瘤以完成增殖并最终释放到土壤中。在自然或实验室条件下获得共生基因的菌株进入到土壤环境后,会面临各种压力。一方面是与土壤中其它微生物的生存竞争;另一方面,菌株需要适应渗透压、营养、水、pH和温度等环境因素的变化[43]。许多实验室条件下构建的菌株虽然能够侵染和诱导相应豆科植物形成瘤状突起,但不能发挥共生固氮作用[3]。对于这种只消耗宿主植物营养而不提供氮源的菌株,植物会通过宿主制裁、免疫系统抑制、养分控制等方式来抑制其繁殖[44]。面对来自植物和土壤环境的胁迫,细菌会通过提高自身突变率等方式进行应对。在大多数根瘤菌的基因组或共生质粒上都含有imuABC基因簇,可以合成易错DNA聚合酶,imuABC的表达能够使细菌发生高频突变[45]。有研究表明当共生基因与编码应激反应的DNA聚合酶基因共同转移时,可以提高受体基因组的突变率从而加速土壤细菌进化为根瘤菌的进程[45]。通过腐生-共生的转换过程可以确保对植物宿主有益的突变株能够得到保留并逐步进化[24]。
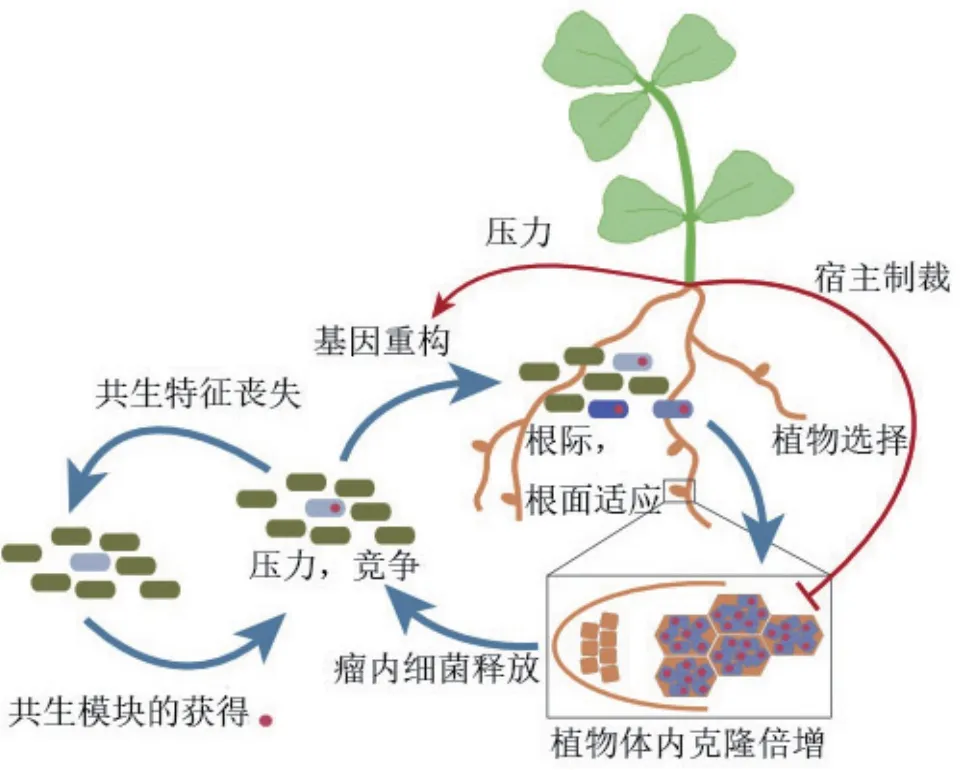
图2 土壤细菌通过基因水平转移获得共生岛后所面临的选择压力[24]
6 展望
根瘤菌作为一类能够与大多数豆科植物进行共生固氮的益生细菌,其宿主专一性问题限制了其应用范围,长期以来研究人员开展了大量的工作,但效果并不明显。近年来关于根瘤菌基因水平转移的研究为解决这一问题提供了新思路。随着合成生物学的发展,以根瘤菌的共生质粒或共生岛为共生固氮元件,通过优化改造这些固氮元件,并向植物根际中其它非共生益生细菌进行转移,从而构建出具有新的共生固氮回路的菌株,并通过实验进化的手段提高菌株的宿主适应性。因为结瘤基因簇的同源性较高,不同专一性的根瘤菌间差异不大,可以将不同专一性的结瘤基因组合成新的结瘤元件进行转移,这样有可能获得新的广宿主根瘤菌菌株。这项工作对于筛选出环境适应性强的新型根瘤菌菌株,扩大根瘤菌种质资源和宿主范围,并最终应用于非豆科植物共生固氮体系建立具有重要的意义。
猜你喜欢
语文世界(小学版)(2022年2期)2022-06-30 14:20:40
山东冶金(2022年1期)2022-04-19 13:40:14
上海金属(2021年4期)2021-07-28 12:33:22
园艺与种苗(2021年1期)2021-03-16 03:01:08
上海金属(2020年6期)2021-01-04 12:25:52
西藏农业科技(2019年3期)2019-11-04 00:35:08
广西林业科学(2016年2期)2016-03-20 05:53:24
——固氮微生物
新疆农垦科技(2016年3期)2016-02-20 23:26:18
河南科技(2014年8期)2014-02-27 14:07:58
植物营养与肥料学报(2012年5期)2012-10-26 03:28:48
