
117份小麦种质中5个矮秆基因的分子检测
2019-10-22 02:25陈向东吴晓军胡喜贵姜小苓李笑慧李小利胡铁柱茹振钢
新疆农业科学 2019年8期
陈向东,吴晓军,胡喜贵,李 淦,姜小苓 ,李笑慧,李小利,胡铁柱,茹振钢
(河南科技学院小麦中心/河南省杂交小麦重点实验室/现代生物育种河南省协同创新中心,河南新乡 453003)
0 引 言
【研究意义】自20世纪60年代以来,以矮秆品种推广和配套栽培方式提高了主要农作物的产量[1]。小麦矮秆种质资源是育成小麦矮秆品种的材料基础,分析小麦种质中矮秆基因分布情况,发掘和鉴定新的优良矮秆基因,创制新种质,对提高小麦育种水平具有重要意义。【前人研究进展】目前,已命名并统一编号的普通小麦Rht矮秆基因有24个。这些矮秆基因中9个来源于自然变异,13个来源于物理化学诱变[2-5]。其中,在我国小麦育种中应用最广泛的有来源于日本农林10号的Rht-B1b(Rht1)、Rht-D1b(Rht2)和日本赤小麦的Rht8、Rht9。另外,来自中国西藏地方品种大拇指矮的Rht-B1c(Rht3)和西安市农科所从“矮秆早”中选出的天然突变系矮变1号的Rht-D1c(Rht10)也有一定的育种应用。Rht基因一般是多效基因,不仅能够降低小麦植株高度,而且有些还能影响产量等农艺性状。Rht-B1b、Rht-D1b基因能够显著降低植株高度,当两者同时存在时,降秆效应还能够累加[6-7]。另外,两者对提高小麦产量具有促进作用[8-9]。一方面表现为直接效应:Rht-B1b能够提高单株的分蘖成穗率、单株粒数、单株粒重等,Rht-D1b能够提高分蘖成穗率、穗粒数、经济系数等;一方面表现为间接效应:Rht-B1b能够增加旗叶和倒二叶面积,增强茎秆强度,Rht-D1b抗倒能力突出,补偿了生物产量降低对产量的负面影响。Rht-B1c和Rht-D1c对产量具有负效作用,如生物产量降低、生长发育缓慢、易受干热风影响和易早衰等[10-11]。Rht4、Rht5、Rht6 和Rht7 在降低植株高度的同时会大大降低粮食产量。Rht8对结实率、籽粒产量和地上部生物产量均有显著的正向作用,能够明显缩短生育期,对不良环境有较强的适应性,但品质性状表现较差[12]。Rht9在降低植株高度的同时会增加分蘖数,穗变短,但最终增加粮食产量[13]。Rht13是对 GA3敏感的矮秆基因,可以降低株高而不影响胚芽鞘长度和幼苗生长势,同时可以增加穗粒数,在小麦遗传改良中颇具应用潜力[14]。【本研究切入点】目前,小麦矮秆基因的分子标记研究取得了很大进展。1999年Peng等克隆出Rht-B1b和Rht-D1b的同源基因。2002年Ellis等设计出能够对Rht-B1b和Rht-B1a及Rht-D1b和Rht-D1a等位基因精确检测的STS引物。2005年Ellis等对Rht4、Rht5、Rht8、Rht9、Rht12和Rht13等矮秆基因进行精细定位,开发了紧密连锁的分子标记。Korzun等和Worland等1998年发现利用SSR引物Xgwm261扩增得到的特异片段可以检测Rht8基因的有无。这些紧密连锁分子标记的开发对小麦育种中矮秆基因的利用起到了促进作用。近年来一些学者对部分国外、国内品种及种质资源进行了矮秆基因分子检测,但选择的分子标记多集中于Rht-B1b、Rht-D1b和Rht8,对其它矮秆基因发掘和利用较少[15-19]。研究我国小麦主产区部分小麦品种中矮秆基因的分布。【拟解决的关键问题】以我国小麦主产区117份小麦品种及种质为材料,利用矮秆基因分子标记进行检测,分析矮秆基因的分布特点和发掘携带特定矮秆基因的新种质,为小麦育种提供技术支撑和材料基础。
1 材料与方法
1.1 材 料
117份供试材料均由河南科技学院小麦中心提供,包含国内推广品种及中间品系等。其中,小偃22携带Rht-B1b和Rht8基因,郑9023携带Rht-D1b基因,扬麦18含Rht8基因,中国春不含任何矮秆基因。
检测Rht-B1a基因(野生型)所用的引物对为BF-WR1,可以扩增出一条长237 bp的条带,检测Rht-B1b基因(突变型,具有致矮作用)所用的引物对为BF-MR1,也同样可以扩增出一条长237 bp的条带。使用两个引物对检测同一个材料,扩增条带的有无,是互补出现的。试验以昝凯等报道的小偃22携带Rht-B1b和Rht8基因[19]作为对照。试验中检测Rht-D1a和Rht-D1b基因所用引物对分别是DF2-WR2和DF-MR2,两者分别可以扩增出264 bp和254 bp的条带,两个引物对扩增条带的有无也是互补出现的。试验以王玉叶报道的郑9023携带Rht-D1b基因[20]为对照。
试验中检测Rht8基因所用引物为Xgwm261,扩增片段长度为192 bp。试验以王玉叶[20]报道的扬麦18携带Rht-D1b基因为对照。
1.2 方 法
1.2.1 田间株高调查
117份小麦材料连续多年种植于河南科技学院辉县试验田,每个材料种植2行,行距20 cm,株距10 cm,采用常规大田管理。在小麦成熟期测量株高,每个材料随机测5株。
1.2.2 DNA提取
将每个材料5株小麦苗期叶片混合,采用CTAB法提取小麦基因组DNA,利用微量分光光度计测量DNA质量和浓度,并将DNA原液稀释成50 ng/μL工作液,放至4℃冰箱保存备用。
1.2.3 PCR扩增
所用矮秆基因检测引物均由上海Invitrogen英俊生物技术有限公司合成,引物序列、退火温度、扩增片段长度等。表1
PCR反应体系:20 μL体系,包含2×TaqMaster Mix(北京康为世纪生物科技有限公司)10 μL,10 μmol/L正反向引物各0.8 μL,50 ng/μL的模板DNA 1 μL,ddH2O 7.4 μL。使用ABI Gene Amp PCR System 9700型PCR仪,扩增程序为95℃预变性 5 min;94℃变性30s,55~63℃退火30s,72℃延伸50s,35循环;72℃延伸5 min。Rht-B1a、Rht-B1b、Rht-D1a和Rht-D1b用1.2%的琼脂糖(含goldview染料)进行电泳检测,Rht8、Rht9、Rht13用2%琼脂糖进行电泳检测,再用凝胶成像系统观察结果并拍照保存。
表1 所用的矮秆基因分子标记
Table 1 Molecular markers of dwarf genes used in this study
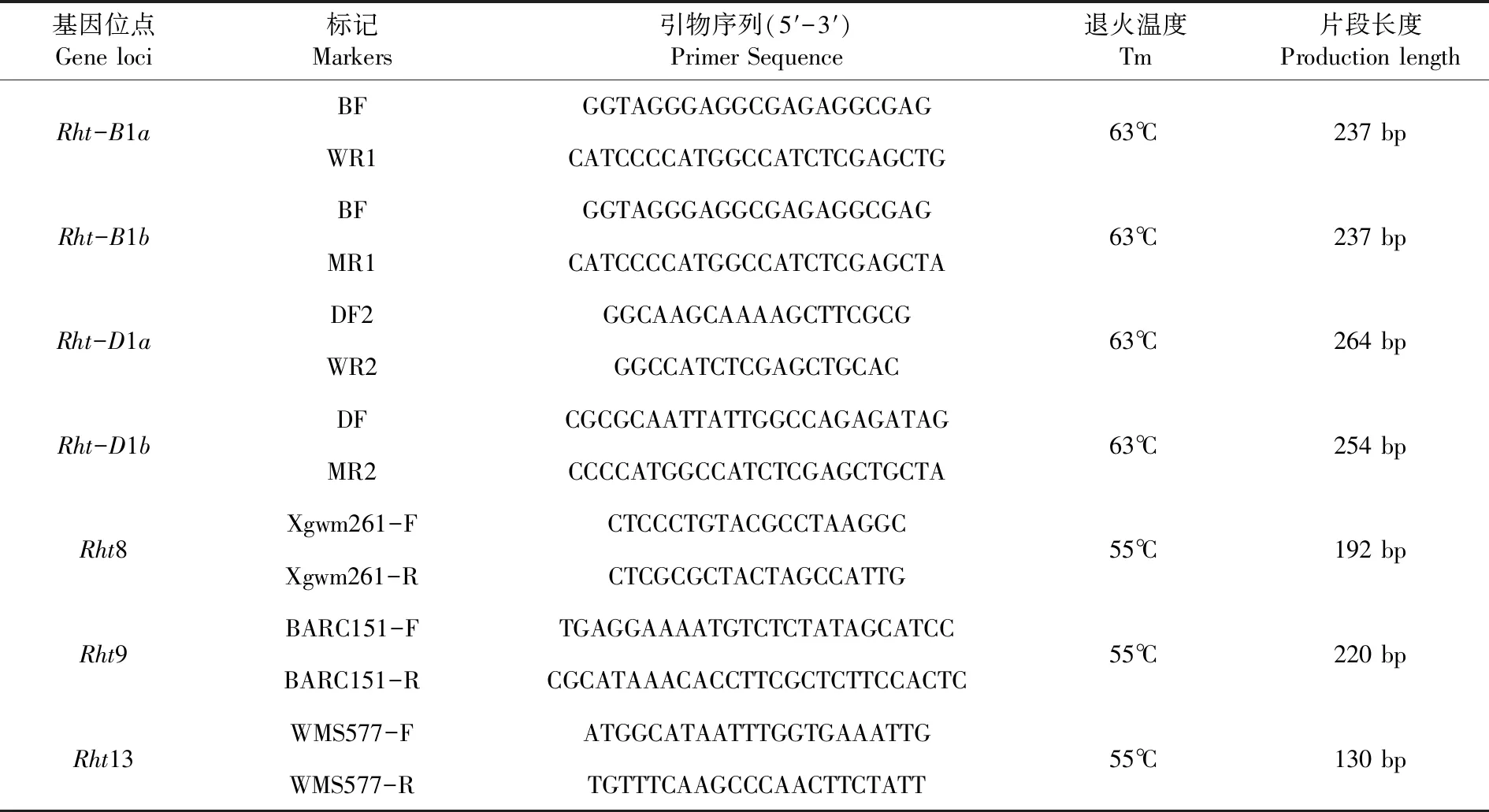
基因位点Gene loci标记Markers引物序列(5′-3′)Primer Sequence退火温度Tm片段长度Production lengthRht-B1aBFGGTAGGGAGGCGAGAGGCGAGWR1CATCCCCATGGCCATCTCGAGCTG63℃237 bpRht-B1bBFGGTAGGGAGGCGAGAGGCGAGMR1CATCCCCATGGCCATCTCGAGCTA63℃237 bpRht-D1aDF2GGCAAGCAAAAGCTTCGCGWR2GGCCATCTCGAGCTGCAC63℃264 bpRht-D1bDFCGCGCAATTATTGGCCAGAGATAGMR2CCCCATGGCCATCTCGAGCTGCTA63℃254 bpRht8Xgwm261-FCTCCCTGTACGCCTAAGGCXgwm261-RCTCGCGCTACTAGCCATTG55℃192 bpRht9BARC151-FTGAGGAAAATGTCTCTATAGCATCCBARC151-RCGCATAAACACCTTCGCTCTTCCACTC55℃220 bpRht13WMS577-FATGGCATAATTTGGTGAAATTGWMS577-RTGTTTCAAGCCCAACTTCTATT55℃130 bp
2 结果与分析
2.1 矮秆基因Rht-B1b检测
研究表明,117份材料中,56份材料携带Rht-B1b基因,占47.9%,平均株高为87.3 cm,变异范围为52~110 cm。只考虑在我国育种应用最广的Rht-B1b、Rht-D1b和Rht8三种矮秆基因,在117份材料中,有Rht-B1b基因而无Rht-D1b和Rht8基因的材料有21份,平均株高88.1 cm,变异范围63~105 cm。表2,图1
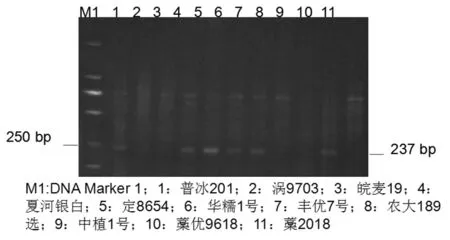
图1 部分材料Rht-B1b基因检测结果
Fig.1 Detection results ofRht-B1bgene
表2Rht-B1b、Rht-D1b和Rht8三种矮秆基因及其组合株高
Table 2 Plant height traits ofRht-B1b,Rht-D1bandRht-8 dwarf genes

基因组成Genes材料(份)Cutivars(lines)平均株高(cm)Plant height变异范围(cm)Variation rangeRht-B1b2188.163~105Rht-D1b882.565~100Rht81889.275~117Rht-B1b、Rht-D1b485.576~100Rht-B1b、Rht81690.774~110Rht-D1b、Rht81984.174~115Rht-B1b、Rht-D1b、Rht81583.274~98
2.2 矮秆基因Rht-D1b检测
研究表明,117份材料中,46份材料携带Rht-D1b基因,占39.3%,平均株高为83.7 cm,变异范围为52~115 cm。如只考虑Rht-D1b、Rht-D1b和Rht8三种矮秆基因,在117份材料中,有Rht-D1b基因而无Rht-B1b和Rht8基因的材料有8份,平均株高82.5 cm,变异范围65~100 cm。表2,图2

图2 部分材料Rht-D1b基因检测
Fig.2 Detection results ofRht-D1bgene
2.3 矮秆基因Rht8的检测结果
研究表明,117份材料中,68份材料携带Rht8基因,占58.1%,平均株高为86.8 cm,变异范围为52~115 cm。只考虑Rht-D1b、Rht-D1b和Rht8三种矮秆基因,在117份材料中,有Rht8基因而无Rht-B1b和Rht-D1b基因的材料有18份,平均株高89.2 cm,变异范围75~117 cm。表2,图3
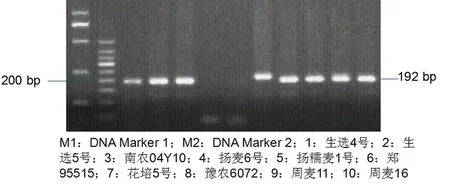
图3 部分材料Rht8基因检测
Fig.3 Detection results ofRht8 gene
2.4 矮秆基因Rht-D1b、Rht-D1b和Rht8的多组合检测
一些小麦品种中Rht-D1b、Rht-D1b和Rht8三种矮秆基因组合出现,117份材料中,同时携带Rht-B1b、Rht-D1b基因的材料有4份,占3.4%,平均株高为85.5 cm,变异范围为76~100 cm;同时携带Rht-B1b、Rht8基因的材料有16份,占13.7%,平均株高为90.7 cm,变异范围为74~110 cm;同时携带Rht-D1b、Rht8基因的材料有19份,占16.2%,平均株高为84.1 cm,变异范围为74~115 cm;同时携带Rht-B1b、Rht-D1b和Rht8基因的材料有15份,占12.8%,平均株高为83.2 cm,变异范围为74~98 cm。表2
2.5 矮秆基因Rht9和Rht13的检测
试验中检测Rht9基因所用引物对为BARC151-F和BARC151-R,扩增片段长度为220 bp。研究表明,117份材料中,18份材料携带Rht9基因,占15.4%,平均株高为92.6 cm,变异范围为65~117 cm。检测Rht13基因所用引物对为WMS577-F和WMS577-R,扩增片段长度为130 bp。117份材料中,14份材料携带Rht13基因,占12.0%,平均株高为93.4 cm,变异范围为82~117 cm。图4~5,表3
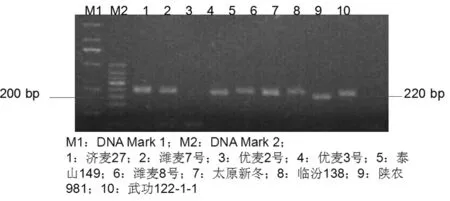
图4 部分材料Rht9基因检测
Fig.4 Detection results ofRht9 gene

图5 部分材料Rht13基因检测
Fig.5 Detection results ofRht13 gene
2.6 117份小麦种质的矮秆基因分布频率分析
研究表明,Rht-B1b、Rht-D1b、Rht8、Rht9和Rht13矮秆基因在不同小麦种质中的分布频率不同,基因组合方式也不一样。只含单个矮秆基因的分布频率高低依次为:Rht-B1b(12,10.3%)>Rht8(13,11.1%)>Rht-D1b(5,4.3%)>Rht13(1,0.9%)>Rht9(0,0.0%)。117份种质中未发现Rht9基因单独携带的情况,而以基因组合形式出现的有18份种质;单独携带Rht13基因的情况也较少,只有1份种质,以基因组合形式出现的有14份种质。5个矮秆基因以同时携带2个基因的组合形式出现,占到37.8%,分布频率高低依次为:Rht-D1b、Rht8(15,12.8%)>Rht-B1b、Rht8(12,10.3%)>Rht-B1b、Rht9(5,4.3%)>Rht-B1b、Rht-D1b(3,2.6%)=Rht8、Rht13(3,2.6%)>Rht-B1b、Rht13(2,1.7%)=Rht-D1b、Rht9(2,1.7%)>Rht-D1b、Rht13(1,0.9%)=Rht8、Rht9(1,0.9%)>Rht9、Rht13(0,0)。5个矮秆基因以同时携带3个基因及以上的组合形式出现,分布频率为21.4%;其中以同时携带Rht-B1b、Rht-D1b和Rht8基因种质最多,占到12.8%。另外,5种矮秆基因均不携带的种质有15份,占到12.8%。表3
表3 我国小麦主产区117份种质的矮秆基因类型及株高
Table 3 Dwarf gene types and plant height of 117 germplasms from main wheat production areas in China
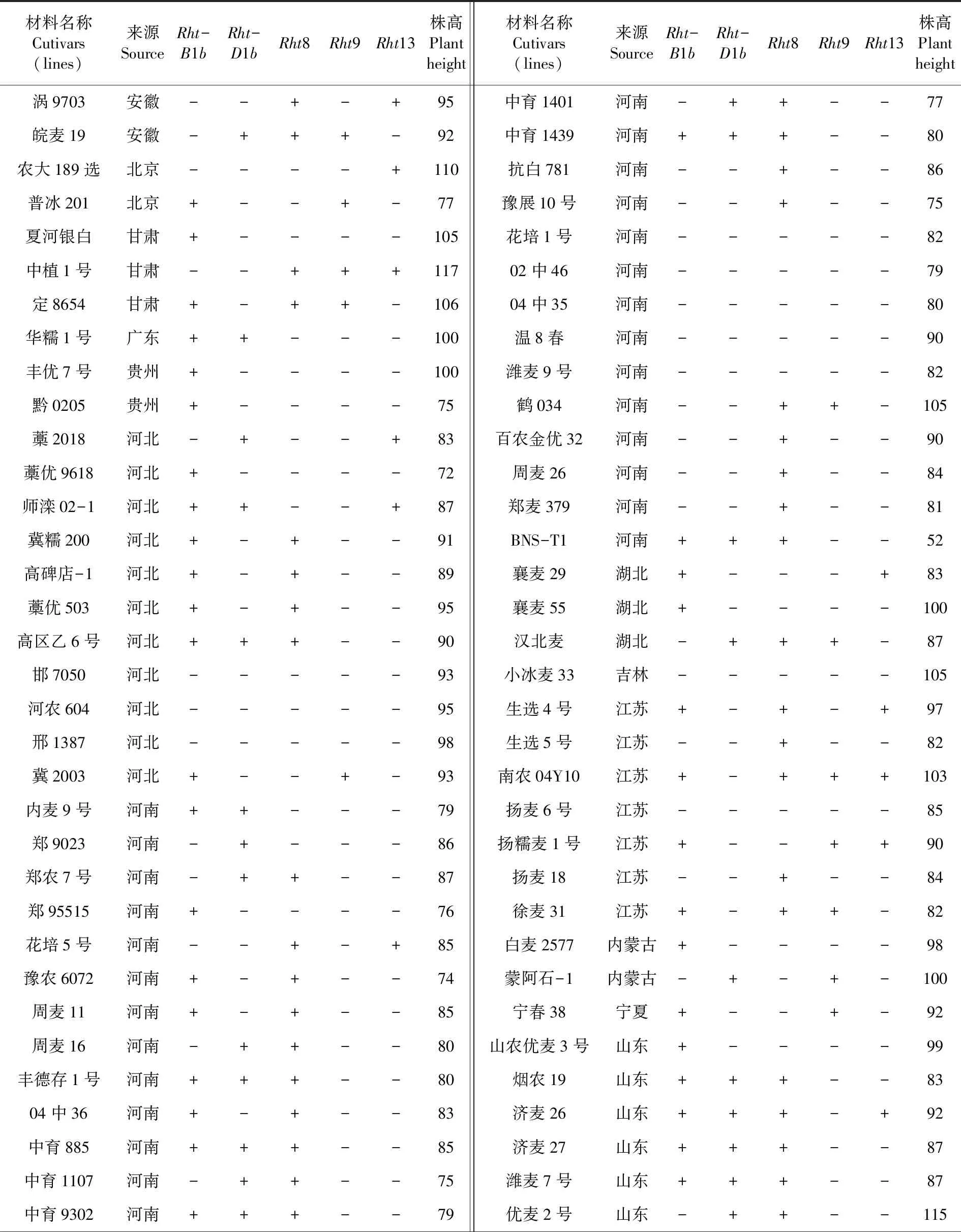
材料名称Cutivars(lines)来源SourceRht-B1bRht- D1bRht8Rht9Rht13株高Plant height材料名称Cutivars(lines)来源SourceRht-B1bRht- D1bRht8Rht9Rht13株高Plant height涡9703安徽--+-+95中育1401河南-++--77皖麦19安徽-+++-92中育1439河南+++--80农大189选北京----+110抗白781河南--+--86普冰201北京+--+-77豫展10号河南--+--75夏河银白甘肃+----105花培1号河南-----82中植1号甘肃--+++11702中46河南-----79定8654甘肃+-++-10604中35河南-----80华糯1号广东++---100温8春河南-----90丰优7号贵州+----100潍麦9号河南-----82黔0205贵州+----75鹤034河南--++-105藁2018河北-+--+83百农金优32河南--+--90藁优9618河北+----72周麦26河南--+--84师滦02-1河北++--+87郑麦379河南--+--81冀糯200河北+-+--91BNS-T1河南+++--52高碑店-1河北+-+--89襄麦29湖北+---+83藁优503河北+-+--95襄麦55湖北+----100高区乙6号河北+++--90汉北麦湖北-+++-87邯7050河北-----93小冰麦33吉林-----105河农604河北-----95生选4号江苏+-+-+97邢1387河北-----98生选5号江苏--+--82冀2003河北+--+-93南农04Y10江苏+-+++103内麦9号河南++---79扬麦6号江苏-----85郑9023河南-+---86扬糯麦1号江苏+--++90郑农7号河南-++--87扬麦18江苏--+--84郑95515河南+----76徐麦31 江苏+-++-82花培5号河南--+-+85白麦2577内蒙古+----98豫农6072河南+-+--74蒙阿石-1内蒙古-+-+-100周麦11河南+-+--85宁春38宁夏+--+-92周麦16河南-++--80山农优麦3号山东+----99丰德存1号河南+++--80烟农19山东+++--8304中36河南+-+--83济麦26山东+++-+92中育885河南+++--85济麦27山东+++--87中育1107河南-++--75潍麦7号山东+++--87中育9302河南+++--79优麦2号山东-++--115
续表3 我国小麦主产区117份种质的矮秆基因类型及株高
Table 3 Dwarf gene types and plant height of 117 germplasms from main wheat production areas in China

材料名称Cutivars(lines)来源SourceRht-B1bRht- D1bRht8Rht9Rht13株高Plant height材料名称Cutivars(lines)来源SourceRht-B1bRht- D1bRht8Rht9Rht13株高Plant height中育9307河南+----76优麦3号山东+++--9605中27河南-+---90泰山149山东--+--100温麦6号河南-++--79潍麦8号山东-----85矮早781河南+-+--85太原新冬山西--+--103偃师4110河南-++--88临汾138山西+--++85天民矮早八A系河南-+++-74陕农981陕西+++--98兰考198河南-++--75武功122-1-1陕西+----87漯麦16河南-----82小偃22-10纯系陕西+-+--95内乡188河南+++--81抗赤6号陕西+---+98洛麦23河南-++--79矮秆962早陕西+++--84洛麦29河南--+--92陕麦94-1-44陕西-++--75泛麦803河南+++--74B91043陕西+--+-89驻麦6号A河南-----81陕808陕西-+---86济创28号河南++---76川育55871四川-----100内乡201河南-+-+-65川麦107四川-++--100科育818河南-+---95川农19四川--+--80洑麦053河南-++--75川育40755四川--+--83中原008河南-++--75绵阳2000-13四川+-+--84新麦208河南-++--80川农21四川+----86新麦18河南-++--80川农23四川--+-+82平原50河南+-+--110新春9号新疆+--+-105百农矮抗58河南+-+--74云麦53云南+-+--98百农160河南+----63云南麦云南-+++-105荔矮4330-1河南-+---55中国春四川-----121中育1311河南--+--82
3 讨 论
矮秆基因的利用有利于提高小麦的抗倒性和进一步挖掘其产量潜力。由于矮秆基因隐性类型的较多,在小麦育种中早代并不宜选择,而结合分子标记检测,可以明确矮秆基因的有无及类型,最终加快育种进程。
各矮秆基因的降秆能力各不相同,研究表明,携带Rht-B1b植株的降秆幅度达到24%,与分别携带Rht-D1b、Rht9、Rht13植株的降秆能力差异不大,而Rht-B1b与Rht-D1b组合会产生累加作用,降秆幅度能达到60%;携带Rht8的植株相对于它的高秆品种降秆11%左右,属于弱降秆基因[21]。研究对国内小麦主产区的117份种质进行了矮秆基因分子检测,结果表明,5个矮秆基因的降秆效应依次为:Rht-D1b>Rht-B1b>Rht8>Rht9>Rht13;就3种矮秆基因Rht-B1b、Rht-D1b和Rht8的组合而言,其降秆效应依次为:Rht-B1b+Rht-D1b+Rht8>Rht-D1b+Rht8>Rht-B1b+Rht-D1b>Rht-B1b+Rht8;与之前其他学者的研究结果基本一致[22-23]。而在试验中Rht-D1b+Rht8的降秆效应稍大于Rht-B1b+Rht-D1b,分析原因可能是受环境条件影响和小麦种质数量有限造成的。另外,对矮秆基因与株高进行t测验的结果发现Rht-D1b与株高呈极显著正相关,Rht9和Rht13与株高呈极显著负相关,而在研究中Rht9与Rht13基本上都是与其他矮秆基因以组合的形式出现,由此原因可能是Rht9与Rht13与其他矮秆基因的组合产生了负向累加作用。
大多数矮秆基因具有一定的降秆作用,能够提高抗倒伏和具有较高的肥水反应,但是存在一些缺陷,如矮秆基因与不利基因连锁,影响小麦的抗病性、抗逆性、产量,熟期等;部分矮秆基因在农艺性状上表现较差,利用价值有限。目前,生产上利用的品种,其携带矮秆基因多集中于Rht-B1b、Rht-D1b和Rht8基因等少数类型,遗传基础狭窄,因此,寻找新的有应用潜力的矮秆基因十分必要。矮秆基因Rht13是一个部分隐性基因,对赤霉素GA3敏感,其主要通过缩短小麦茎节长度而降低株高,增加小麦产量;与Rht-D1b相比,不影响胚芽鞘的长度和幼苗生长活力,具有较大育种应用价值[24]。
4 结 论
研究对国内小麦主产区的部分品种进行了5个矮秆基因分子标记检测,明确了这些品种所携带的矮秆基因。117份种质材料中,矮秆基因分布频率依次为Rht8(68份,58.1%)>Rht-B1b(56份,47.9%)>Rht-D1b(46份,39.3%)>Rht9(18份,15.4%)>Rht13(14份,12.0%)。部分种质中同时携带2个矮秆基因的组合占到37.8%;同时携带3个及以上基因的组合占到21.4%,其中以同时携带Rht-B1b、Rht-D1b和Rht8基因种质最多,占到12.8%。5种矮秆基因均不携带的种质有15份,占到12.8%。鉴定出14份携带Rht13矮秆基因的新种质。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
中国农业大学学报(2022年3期)2022-05-19
作物学报(2022年3期)2022-01-22
麦类作物学报(2021年5期)2021-09-23
大麦与谷类科学(2020年1期)2020-03-30
四川蚕业(2020年4期)2020-02-10
种子(2019年12期)2020-01-07
农村百事通(2019年17期)2019-10-08
现代农业科技(2017年1期)2017-03-06
