
鹅观草种子萌发及出苗特性
2019-10-21 06:21谢元周伟军陆永良于晓玥唐伟
杂草学报 2019年1期
谢元 周伟军 陆永良 于晓玥 唐伟

摘要:在实验室和温室条件下研究不同温度、光照、水势、盐分对鹅观草种子萌发以及埋土深度对鹅观草出苗的影响。结果表明,鹅观草种子萌发的昼/夜温度范围为20 ℃/10 ℃~35 ℃/25 ℃,最适昼/夜温度为 25 ℃/15 ℃;黑暗条件下,鹅观草种子萌发率较光照下略有提高,说明鹅观草萌发不需要光照;鹅观草种子对水势胁迫较敏感,水势从0下降至 -0.8 MPa 时,发芽率从93%直线下降至0;鹅观草种子对盐分胁迫具有一定耐受性,当NaCl浓度≤50 mmol/L时,种子均能保持80%以上的萌发率。50%萌发抑制率时的水势和NaCl浓度分别约为-0.4 MPa和100 mmol/L。埋土深度为0.5 cm时,鹅观草出苗率最高,埋土深度≥3 cm时出苗率显著下降。表明鹅观草种子萌发具有较强的环境适应性,通过耕作将其种子带入4 cm以下土层,可有效抑制鹅观草的危害。
关键词:鹅观草;种子萌发;种子出苗;环境因子
中图分类号:S451 文献标志码:A 文章编号:1003-935X(2019)01-0017-06
Abstract:Laboratory and greenhouse experiments were conducted to study the effects of temperature,light,osmotic and salt stress on seed germination,and the effect of burial depth on seedling emergence of Roegneria kamoji Ohwi. The temperature range for germination is 20/10 ℃~35/25 ℃,with the optimal day/night temperature 25 ℃/15 ℃. Under dark conditions,germination rate increased slightly,indicating that light was not necessary. Seed germination was sensitive to osmotic stress,and when the water stress dropped from 0 to -0.8 MPa,the germination rate decresedfrom 93% to 0. The seed could tolerate salt stress. When NaCl concentration is lower than 50 mmol/L,seed germinationrate can be maintained at over 80%. The osmotic potential and NaCl concentration required for 50% inhibition of maximum germination were -0.4 MPa and 100 mmol/L,respectively. The seedling emergence rate was highest when the burial depth was 0.5 cm,and it was significantly inhibited when the burial depth was over 4 cm. These results indicated that R. kamojihad relatively strong environmental adaptability,potentiallybecoming a competitive weed in wheat field. Soil inversion by tillage to bury weed seeds below their maximum depth of emergence may enable effective control of this weed.
Key words:Roegneria kamoji Ohwi;seed germination;seedlin gemergence;environmental factors
鵝观草(Roegneria kamoji Ohwi)(2n=6x=42)是小麦族禾本科鹅观草属的一种六倍体多年生禾草,拥有耐湿、高抗赤霉病等优良品性,是小麦近缘植物改良和牧草育种的天然基因库[1-2]。营养价值高、适口性好等优点使其成为优良牧草,同时鹅观草还具有生态功能,可用于改善生态环境[3-4]。鹅观草多生长在海拔100~2 300 m的山坡和湿润草地上,除青海、西藏等地外,其分布遍及全国。鹅观草在农田中偶有发现,危害不重,但目前在个别地区已成为农田的主要杂草,且危害程度不断上升[5-6]。目前,主要利用形态学、细胞学、同工酶、随机扩增多态性DNA标记(RAPD)、基因组原位杂交(GISH)等手段对小麦族属种间、鹅观草属种间的遗传变异进行研究,而对鹅观草种子萌发及出苗特性的研究未见系统报道。本试验研究环境因子对鹅观草种子萌发和出苗的影响,以期为今后鹅观草的防除工作提供理论依据。
1 材料与方法
1.1 材料
鹅观草种子于2017年5月采自湖北省荆州市(30°13′N,112°33′E)小麦田,将采自50株植株上的完全成熟的种子装于纸袋中,风干后于20 ℃左右的实验室保存4个月,备用。
1.2 方法
选取籽粒饱满、无病变的种子,用0.1% HgCl2溶液消毒3 min后,用无菌水充分冲洗干净,晾干备用[7]。关于鹅观草萌发对不同温度、光照、水势、盐分响应的研究在培养皿中进行。在直径为 9 cm 的玻璃培养皿中铺垫2层滤纸,并向每个培养皿中加入5 mL配制好的测试溶液或蒸馏水,每个处理放置25粒种子,重复3次,试验期间每天补充因蒸发而散失的水分。将进行温度、光照、水势、盐分处理试验的培养皿放置于光照度为 3 000 lx 的培养箱内培养[8-9]。关于埋土深度对鹅观草出苗影响的研究在塑料钵中进行。
1.2.1 温度对萌发的影响 设置5个温度处理,即光照/黑暗条件下的温度为15 ℃/5 ℃、20 ℃/10 ℃、25 ℃/15 ℃、30 ℃/20 ℃、35 ℃/25 ℃等处理,光照—黑暗周期为12 h—12 h。每天定时观察并记录培养皿内种子的萌发情况,连续观察15 d[10]。
1.2.2 光照对萌发的影响 在“1.2.1”节不同温度处理的基础上,再分别对应设置1个完全黑暗条件处理,完全黑暗处理同样设置3次重复,用不透光的黑色遮光布遮盖以制造完全黑暗的条件,其他条件相同。试验期间保持黑暗条件,第15天除去遮光布,记录培养皿内种子的萌发情况[11]。
1.2.3 水势胁迫对萌发的影响 参考Michel等关于溶液浓度与渗透势的计算方法[12],使用聚乙二醇(PEG-8000)配制渗透势为0、-0.1、-0.2、-0.3、-0.4、-0.6、-0.8 MPa的不同测试溶液。将各处理置于光照/黑暗温度为25 ℃/15 ℃、光照—黑暗周期为12 h—12 h的培养箱中培养。每天定时观察记录培养皿内种子的萌发情况,连续观察15 d。
1.2.4 盐分胁迫对萌发的影响 采用不同浓度(0、25、50、75、100、125、150 mmol/L)的NaCl溶液测定种子萌发对盐分胁迫的响应。将各处理置于光照/黑暗温度为25 ℃/15 ℃、光照—黑暗周期为12 h—12 h的培养箱中培养[13-15]。每天定时观察记录培养皿内种子的萌发情况,连续观察15 d。
1.2.5 埋土深度对出苗的影响 将鹅观草种子播种于口径为12 cm、高为10 cm、底部有排水孔的塑料钵中,塑料钵底部盖有单层滤纸,钵中装有适量事先高压灭菌的试验用土,底部浸水直至土壤湿润。每钵播种鹅观草种子25粒,播种深度设置为0、0.5、1.0、2.0、3.0、4.0 cm,每个处理重复3次。试验在室外网室中进行,平均气温为20 ℃,试验期间定时补充水分,以保证土壤湿度适宜,每天定时观察记录种子的出苗动态,连续观察20 d。设置没有播种的塑料钵作为空白对照,若20 d后空白对照没有鹅观草出苗,说明试验用土不含鹅观草种子。
1.3 数据处理与分析
在萌发试验中将长出芽长和种子等长视为萌发,在出苗试验中将子叶露出土壤表层视为出苗,并将连续7 d无种子萌发视为萌发结束。根据观察记录的数据计算发芽率、平均发芽时间、发芽启动时间。
发芽率=已发芽种子数/供试种子总数×100%。
平均发芽时间=∑(Ni×Di)/∑Ni。
式中:Ni表示第i天萌发的种子数,粒;Di表示相应的萌发时间。
发芽启动时间为第1粒种子发芽所需时间(d)。
使用Excel进行数据统计,用SPSS 20.0进行方差分析,用Duncans法进行多重比较。
2 结果与分析
2.1 温度对萌发的影响
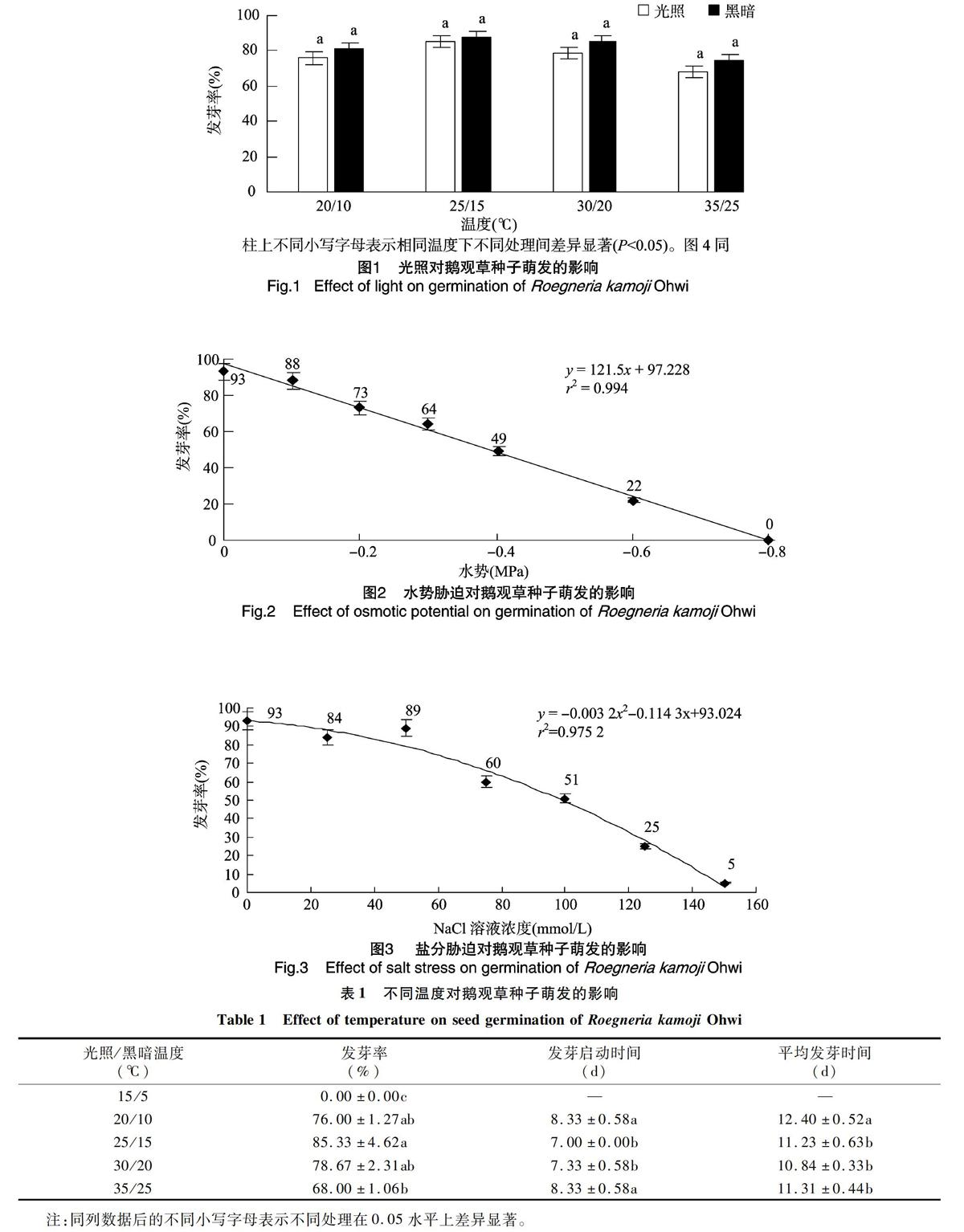
由表1可知,鹅观草种子在光照/黑暗温度为 25 ℃/15 ℃ 时,发芽率最高,可达到85.33%;光照/黑暗温度为20 ℃/10 ℃、30 ℃/20 ℃时,发芽率略有下降,但与25 ℃/15 ℃时的发芽率差异不显著。当光照/黑暗温度升至35 ℃/25 ℃时,发芽率降至68.00%,与 25 ℃/15 ℃ 时的发芽率存在显著差异。当光照/黑暗温度降至15 ℃/5 ℃时,种子不再萌发。从发芽启动时间看,光照/黑暗温度为 25 ℃/15 ℃时最快,但与 30 ℃/20 ℃ 之间不存在显著差异,35 ℃/25 ℃与20 ℃/10 ℃时的发芽启动时间大致相同。从平均发芽时间来看,光照/黑暗温度为30 ℃/20 ℃时的平均发芽时间最短,但与25 ℃/15 ℃、35 ℃/25 ℃之间不存在显著差异,光照/黑暗温度为20 ℃/10 ℃时的平均发芽时间与其他3个温度处理相比显著增长。
2.2 光照对萌发的影响
由图1可知,在相同温度条件下,光照与黑暗处理的鹅观草发芽率没有显著差异,除15 ℃/5 ℃的光照/黑暗温度下均不萌发外,其他温度条件下黑暗处理的发芽率均略高于光照處理,但无显著差异,说明鹅观草种子萌发不需要光照,因此光照不能成为鹅观草田间萌发的影响条件。
2.3 水势胁迫对萌发的影响
由图2可知,当溶液水势从0下降到-0.8 MPa时,鹅观草的发芽率从93%迅速下降至0。发芽率与水势胁迫之间的关系符合线性方程y=121.500x+97.228(r2=0.994)。当水势低至 -0.8 MPa 时,鹅观草种子不再萌发。表明鹅观草种子对水势胁迫较敏感,湿润环境有利于萌发,干旱环境抑制萌发。
2.4 盐分胁迫对萌发的影响
由图3可知,当NaCl溶液浓度从0上升到150 mmol/L时,鹅观草的发芽率从93%降至5%。发芽率与盐分胁迫之间的关系符合方程y=-0.003 2x2-0.114 3x+93.024 0(r2=0.975 2)。当盐分浓度≤50 mmol/L时,鹅观草的发芽率均大于80%;随着盐分浓度的上升,NaCl对鹅观草萌发的抑制作用逐渐增强;当NaCl浓度达到 150 mmol/L 时, 种子几乎不萌发。 表明鹅观草种子对盐分胁迫具有一定的耐受能力,低盐条件下对种子萌发的影响较小。
2.5 埋土深度对出苗的影响
由图4可知,在0、0.5、1.0、2.0、3.0、4.0 cm等6个埋土深度处理中,以埋土深度为0.5 cm的鹅观草种子出苗率最高,但与播于土表的鹅观草出苗率之间不存在显著差异;当播种深度从0增加至2 cm时,鹅观草种子出苗率整体下降,鹅观草在播种深度为2 cm时,仍有76%的高出苗率,推测可能与其萌发不需要光照有关;随着埋土深度的进一步增加,出苗率迅速下降,埋土深度为 0.5 cm 与埋土深度为 2 cm 之间出现显著差异,当埋土深度达到4 cm时,出苗率仅为28%,出苗率显著下降可能与其种子体积小,营养物质不足以提供其出苗所需能量有关。表明鹅观草可以正常萌发的埋土深度范围较广,在下茬作物播种前对
土壤进行深耕,可有效控制鹅观草的危害。
3 结论与讨论
鹅观草种子对水势胁迫比较敏感,随着水势的降低,种子萌发率呈直线下降态势,说明水分充足更有利于鹅观草种子萌发,这與鹅观草多生于湿润环境的特性相符。除此之外,鹅观草的萌发对光照、温度、盐分胁迫和埋土深度等环境因子都有一定的耐受性,适应范围较广,其中光照/黑暗气温低至 15 ℃/5 ℃ 时,种子萌发才受抑制,其他温度均保持较高萌发率,适宜萌发温度较菵草(Beckmannia syzigachne)、看麦娘(Alopecurus aequalis)等麦田杂草更广泛。饶娜报道,菵草种子萌发的最适温度范围是8~16 ℃,高于25 ℃时不能萌发[16],而鹅观草在光照/黑暗温度为35 ℃/25 ℃ 时依然保持较高的萌发率。鹅观草萌发不需要光照,在有无光照的条件下均能保持高萌发率,与菵草、日本看麦娘(Alopecurus japonicus)[17-18]具有相似特性。鹅观草种子对水势胁迫较敏感,湿润环境有利于萌发,干旱环境抑制萌发,这与鹅观草多见于田边、沟渠旁的特性吻合。当NaCl溶液浓度达到100 mmol/L时,鹅观草仍有50%以上的萌发率,表明其耐盐碱能力较强,这与菵草的表现[19]是一致的。相对于猪殃殃(Galium aparine)[20]、野燕麦(Avena fatua)[21]等麦田杂草种子土表不萌发的情况,鹅观草种子大小适中,表面无硬实颖壳,吸水性良好,土表种子也能萌发,同时其胚芽顶土能力较强,萌发土层范围广,当埋土深度达到4 cm时,才能有效抑制其出苗。
鹅观草种子萌发较强的环境适应性有助于其在小麦种植区生存繁殖,在我国长江中下游部分省(市、自治区)已经出现鹅观草进入麦田造成小麦减产的情况,且草害逐年加重,除草剂抗药性逐渐显现,应当引起足够重视。根据鹅观草种子萌发及出苗对温度、光照、水势、盐分和埋土深度的要求,可以指导以鹅观草为优势种杂草群的农田防除工作。除采取人工拔除和化学防除等方法外,根据鹅观草是多年生杂草的特性,在下茬作物播种前,通过翻耕破坏鹅观草根系,同时将种子带入 4 cm 以下土层,从而有效抑制鹅观草的萌发和出苗,控制其危害程度。
参考文献:
[1]蔡联炳. 鹅观草属的地理分布[J]. 西北植物学报,2002,22(4):913-923.
[2]陈志石,吴竞仑,李永丰,等. 麦田土壤杂草种子库研究[J]. 江苏农业学报,2006,22(4):401-404.
[3]肖海峻,翟利剑,卢宏双,等. 小麦族鹅观草属植物研究进展[J]. 草业科学,2007,24(4):41-46.
[4]蔡联炳,王玉金. 鹅观草属的系统发育分析[J]. 植物研究,2001,21(4):554-560.
[5]周 凯,陈仕勇,马 啸,等. 鹅观草属系统演化研究进展[J]. 草业科学,2012,29(9):1457-1461.
[6]Tang W,Chen J,Zhang J P,et al. Seed germination,seedling emergence,and response to herbicides of triquetrous murdannia (Murdannia triquetra) in rice[J]. Weed Science.2017,65(1):141-150.
[7]解继红,于靖怡,陈景芋,等. 鹅观草属种子消毒及无菌播种的研究[J]. 种子,2009,28(3):25-27.
[8]罗小娟,吕 波,李 俊,等. 鳢肠种子萌发及出苗条件的研究[J]. 南京农业大学学报,2012,35(2):71-75.
[9]李 俊,饶 娜,董立尧,等. 稻茬麦田菵草发生动态及其对小麦生长和产量的影响[J]. 南京农业大学学报,2010,33(3):67-70.
[10]周娟娟,尹国丽,魏 巍,等. 两种鹅观草种子萌发对温度和水分的响应[J]. 中国草地学报,2014,36(2):98-103.
[11]解继红,于靖怡,徐 柱,等. 浸种和光照处理对中间鹅观草种子萌发的影响[J]. 中国种业,2009(5):43-44.
[12]Michel B E,Kaufmann M R. The osmotic potential of polyethylene glycol 6000[J]. Plant Physiology,1973,51(5):914-916.
[13]张谨华,郭平毅. 3种鹅观草耐盐鉴定指标的筛选研究[J]. 基因组学与应用生物学,2014,33(6):1341-1345.
[14]解继红,德 英,穆怀彬,等. NaCl胁迫对鹅观草苗期生理生化物质的影响[J]. 中国草地学报,2013,35(5):152-155.
[15]陈景芋,解继红,徐 柱,等. NaCl胁迫对3种鹅观草幼苗耐盐性研究[J]. 中国农学通报,2011,27(1):383-388.
[16]饶 娜. 菵草[Beckmannia syzigachne (Steud.) Fernald]生物学、生态学及其防除的研究[D]. 南京:南京农业大学,2008.
[17]於乐瑞,李 斌. 环境因子对日本看麦娘种子萌发的影响[J]. 大麦与谷类科学,2018,35(1):38-42.
[18]沈国辉,管丽琴,石 鑫,等. 日本看麦娘生物学、生态学特性[J]. 上海农业学报,2000,16(增刊1):37-40.
[19]Rao N,Dong L Y,Li J,et al. Influence of environmental factors on seed germination and seedling emergence of American sloughgrass (Beckmannia syzigachne) [J]. Weed Science.2008,56(4):529-533.
[20]张田田,马 冲,吴翠霞,等. 不同环境因素对猪殃殃种子萌发及出苗的影响[J]. 植物保护,2017,43(2):102-106.
[21]李 涛,袁国徽,钱振官,等. 野燕麦种子萌发特性及化学防除药剂筛选[J]. 植物保护,2018,44(3):111-116.
猜你喜欢
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
天津农业科学(2016年12期)2017-01-11
现代农业科技(2016年20期)2016-12-20
中国市场(2016年36期)2016-10-19
天津农业科学(2015年12期)2015-12-03
河北渔业(2015年10期)2015-10-15
