
叉头样转录因子3和白细胞介素10在慢性根尖周炎组织中的表达研究
2019-10-16 06:38吕广应刘乙臻张秀梅王琳源
实用口腔医学杂志 2019年5期
吕广应 刘乙臻 张秀梅 王琳源
慢性根尖周炎是外界物理、化学和生物性损伤刺激根尖周组织而引起的慢性炎症反应,不仅损伤根尖周组织,还与心血管疾病、糖尿病等系统性疾病密切相关。目前临床上治疗慢性根尖周炎主要是采用根管治疗,但有部分根尖周炎的患者在经过治疗后根尖周组织仍存在炎症,无法从根本上治疗根尖周炎[1-2]。因此,研究慢性根尖炎发生、发展的关键机制,寻找关键的阻断靶点是目前亟待解决的问题。研究发现宿主对抗根尖周致病菌的免疫应答在慢性根尖周炎的发病过程中具有重要作用[3],其中调节性T(regulatory T,Treg)细胞越来越受到人们的关注[4]。Treg细胞特异性地表达叉头翼螺旋转录分子(forhead box P3,Foxp3)和表面分子CD25、细胞毒性T淋巴细胞相关抗原-4(cytotoxic T lymphocyte associated antigen-4,CTLA-4)和糖皮质激素诱导的肿瘤坏死因子受体(glucocorticoid induced tumor necrosis factor receptor,GITR),分泌抑炎性细胞因子白细胞介素10(interleukin-10,IL-10)、转化生长因子β(transforming growth factor-β,TGF-β),在慢性牙周炎、口腔扁平苔藓病损、慢性胃炎等感染性疾病以及多发性肌炎、类风湿性关节炎等自身免疫性疾病中削弱炎症效应,维持免疫系统的稳定[5-9]。研究发现在慢性牙周炎牙周组织破坏期,牙龈组织中Treg细胞数量减少,相关细胞因子(IL-10和TGF-β)表达降低,而在牙周组织修复期,Treg细胞数量增多,相关细胞因子表达增高[10]。Gar-let等[11]采用抗体阻断的方法阻断Treg细胞表面分子GITR,抑制Treg细胞的功能,加重了伴放线放线杆菌(Aggregatibacter actinomycetemcomitans,Aa)诱导的牙周组织炎症程度和牙槽骨吸收。与慢性牙周炎相似,慢性根尖周炎作为一种细菌感染性疾病,以炎症和牙槽骨破坏为主要特征。然而在慢性根尖周炎的病损组织中Treg细胞表现如何,在其发病过程中可能发挥怎样的作用,目前在国内有关这方面的报道较少。本研究旨在通过检测Treg细胞相关细胞因子IL-10和转录因子Foxp3在慢性根尖周炎病损组织中的表达情况,探讨Treg细胞在慢性根尖周炎的作用,为慢性根尖周炎的免疫学研究提供新的理论基础和实验依据。
1 材料与方法
1.1 主要实验材料
Marker(赛默飞公司,美国);Foxp3抗体、IL-10抗体(Cell Signaling公司,美国);β-actin(北京博奥森生物技术有限公司);羊抗兔二抗(EarthOx,LLC公司,美国);PVDF膜(GE公司,美国);SDS-PAGE凝胶配制试剂盒(上海碧云天公司);反转录试剂盒(南京Vazyme公司);Trizol(Ambion公司,美国);BCA蛋白测定试剂盒、Foxp3引物合成、IL-10引物合成、DEPC水(北京鼎国昌盛公司)。
1.2 样本采集和处理
收集2016-12~2017-05锦州医科大学附属第二医院口腔颌面外科门诊15例依据临床表现和X线片所见诊断为慢性根尖周炎,需要进行根尖外科手术或拔除的患牙,取与牙根尖周区相连的病损软组织作为实验组。收集15例埋伏阻生需要拔除的患牙,取阻生牙冠方覆盖的健康牙龈组织作为对照组[12-14]。慢性根尖周炎纳入标准:影像学证实有根尖周骨质破坏;根尖周有相连的病损组织。患者排除标准:①患者有系统性疾病;②患牙有牙周病变;③患牙为牙周牙髓联合病变;④过去6个月内服用抗生素或非甾体类抗炎药物。获取标本后用液氮速冻后-80℃保存备用。在进行临床试验之前,所有患者签署知情同意书。本研究经锦州医科大学伦理委员会审核批准。
1.3 RT-PCR检测
1.3.1 RNA提取 根据Trizol试剂盒说明书操作。用紫外分光光度计测量RNA浓度测定A值,260 nm/280 nm均在1.8~2.2,记录提取的总RNA浓度,单位为ng/μl。操作完成后,冻存于-80℃冰箱或直接进行PCR反应。
1.3.2 RT-PCR 反应 按照一步法反转录试剂盒(Vazyme)配置25μl反应体系,加入反应体系中RNA的量约为300 ng。反转录得到的cDNA进行PCR扩增。引物设计:Foxp3引物序列:上游引物序列:5'-GTGGCCCGGATGTGAGAAG-3',下游引物序列5'-GGAGCCCTTGTCGGATGATG-3',扩增目的基因片段长度为238 bp。IL-10引物序列:上游引物序:5'-GACTTTAAGGGTTACCTGGGTTG-3',下游引物序列5'-TCACATGCGCCTTGATGTCTG-3',扩增目的基因片段长度为112 bp。以β-actin为内参照物,其上游引物序列:5'-AGCGAGCATCCCCCAAAGTT-3',下游引物序列5'-GGGCACGAAGGCTCATCATT-3',扩增目的基因片段长度为285 bp引物均由北京鼎国昌盛生物科技有限公司合成。反应条件为94℃,5 min;35个PCR循环(94℃,30 s;72℃,1 min);最后72℃延伸7 min。PCR产物在2%琼脂糖凝胶分离,在凝胶成像系统内扫描后测量谱带强度。
1.4 Western-blot检测
取出适量标本,室温融化,加入150μl裂解液,反复吹打以充分裂解细胞,用BCA蛋白试剂盒测定蛋白浓度,经蛋白定量后按20μg/孔行10%聚丙烯酰胺(10%SDS-PAGE)电泳并经湿式转膜、洗膜、封闭后,取一抗稀释液(TBST液)按照1∶1 000稀释Foxp3,IL-10抗体,4℃摇床过夜;洗膜后,再加入按照1∶10 000稀释的二抗,室温振摇孵育2 h,充分洗膜后,用ECL法显色、曝光后测量X光胶片上条带密度。
1.5 统计学分析
采用SPSS 17.0软件包对数据进行统计学分析,数据为计量资料,以±s表示,两组间比较采用两独立样本t检验。P<0.05为差异有统计学意义。
2 结 果
2.1 RT-PCR结果
通过RT-PCR比较健康对照组和慢性根尖炎组Treg细胞相关因子Foxp3、IL-10 mRNA表达的差异。在健康牙龈组织和慢性根尖周炎病损组织中均有不同程度Foxp3、IL-10 mRNA的表达,并且慢性根尖周炎组Foxp3、IL-10 mRNA的表达水平显著高于健康对照组,2组间差异显著(P<0.05)有统计学意义(图1,表1)。
2.2 Western-blot检测结果
通过Western-blot检测健康牙龈组织和慢性根尖周炎病损组织Foxp3和IL-10蛋白表达情况。在健康牙龈组织和慢性根尖周炎病损组织中均有Foxp3和IL-10蛋白表达。与健康对照组相比,慢性根尖周炎组中Foxp3和IL-10蛋白表达水平增高,2组间表达量差异显著(P<0.05)有统计学意义(图2,表2)。
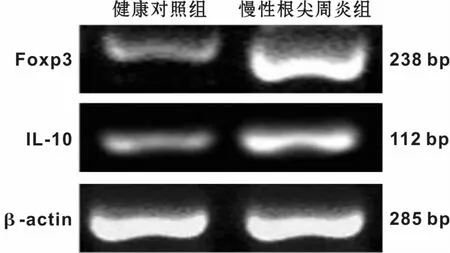
图1 2组中Foxp3、IL-10的mRNA表达Fig 1 The mRNA expression of Foxp3 and IL-10 in the 2 groups
表1 2组中Foxp3、IL-10的mRNA表达及比较 (±s)Tab 1 The mRNA expression of Foxp3 and IL-10 in the 2 groups(±s)
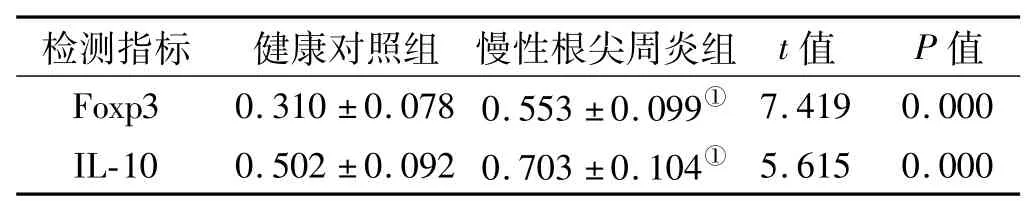
表1 2组中Foxp3、IL-10的mRNA表达及比较 (±s)Tab 1 The mRNA expression of Foxp3 and IL-10 in the 2 groups(±s)
注:①与健康对照组相比,P<0.01
检测指标 健康对照组 慢性根尖周炎组 t值 P值Foxp3 0.310±0.078 0.553±0.099①7.419 0.000 IL-10 0.502±0.092 0.703±0.104①5.615 0.000
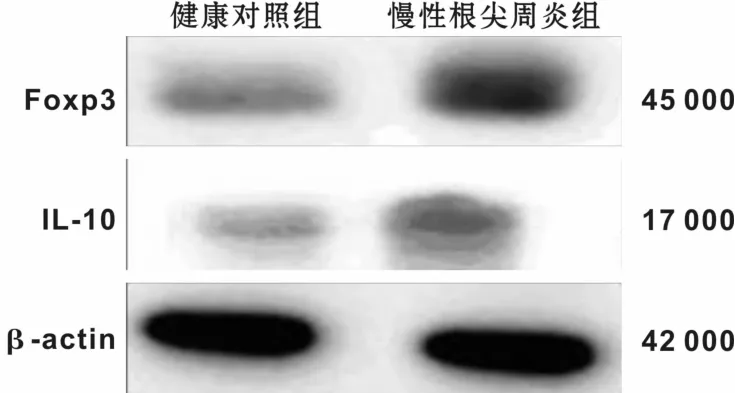
图2 2组中Foxp3、IL-10的蛋白表达Fig 2 The protein expression of Foxp3 and IL-10 in the 2 groups
表2 2组中Foxp3、IL-10的蛋白表达及比较 (±s)Tab 2 The protein expression of Foxp3 and IL-10 in the 2 groups(±s)

表2 2组中Foxp3、IL-10的蛋白表达及比较 (±s)Tab 2 The protein expression of Foxp3 and IL-10 in the 2 groups(±s)
注:①与健康对照组相比,P<0.01
检测指标 健康对照组 慢性根尖周炎组 t值 P值Foxp3 0.319±0.122 0.500±0.143①3.835 0.010 IL-10 0.352±0.162 0.683±0.093①7.086 0.000
3 讨 论
慢性根尖周炎是口腔内科的常见病,表现为炎性肉芽组织的形成和牙槽骨破坏。来自根管系统的细菌感染是慢性根尖周炎主要的致病因素。研究发现病原菌侵入到根尖周组织后,在局部趋化因子的作用下,区域病原菌特异性CD4+T细胞活化、增殖并浸润到病损根尖周组织中在微环境的影响下分化为不同的细胞亚群并发挥抗炎性或促炎性作用[15-16]。
Treg细胞作为CD4+T细胞亚群具有独特的免疫调节功能,通过细胞间直接接触或分泌细胞因子IL-10、TGF-β等方式,抑制体内多种免疫细胞的活化和增殖,进而发挥免疫抑制作用[17]。Foxp3是Treg细胞特异性核转录因子,对Treg细胞的发育、分化及功能的发挥必不可少。清除Treg细胞内的Foxp3则导致Treg细胞功能丧失,并且效应性T细胞功能加强,免疫促进相关因子表达增多[18]。阳莎莎等[19]在研究Foxp3的表达与大鼠实验性根尖周炎骨吸收的关系时发现,在根尖周炎急性期Foxp3的表达水平较低,破骨细胞增多伴随骨吸收加重;而在根尖周炎慢性期Foxp3的表达增高,破骨细胞数目减少且骨吸收无扩大趋势。提示Treg细胞可能在根尖周炎控制炎症进展过程中发挥重要作用。Campos等[20]发现在根尖肉芽肿和根尖囊肿的非活动性病损组织中Foxp3的表达增高,并且与IL-10和TGF-β的表达水平正相关,表明Foxp3调控Treg细胞免疫调节功能(包括免疫抑制和缓解炎症反应)的发挥,提示根尖病损组织中抗炎性细胞因子IL-10和TGF-β可能主要来源于Treg细胞。IL-10作为抗炎性细胞因子,通过抑制过度活化的效应T细胞,以及下调促炎因子的产生调节免疫应答,抑制病变组织炎症发展,抑制骨吸收[21]。Sasaki等[22]在实验性小鼠牙髓感染诱导的根尖骨吸收模型中发现IL-10缺乏小鼠的根尖骨吸收明显加重,体外研究发现IL-10以剂量依赖性方式抑制促炎性细胞因子IL-1β的产生,表明IL-10在抑制感染诱导根尖周骨吸收中发挥重要的作用。Garlet等[11]研究发现当牙周组织发生破坏时,Treg细胞数量减少,相应的细胞因子(IL-10和TGF-β)表达降低,而在组织修复期,病变恢复期时,病变牙龈组织中出现增多的Treg,并且Treg相关的细胞因子表达上调。与上述研究结果相一致,该研究发现在健康牙龈组织和慢性根尖周炎病损组织中均存在Treg细胞特征性转录因子Foxp3和相关细胞因子IL-10的表达,并且慢性根尖周炎病损组织中Foxp3和IL-10表达显著高于健康牙龈组织,表明在慢性根尖周炎中Treg细胞介导的免疫应答增强,提示Treg细胞参与维持根尖周组织稳态和组织损伤的修复。
此外,Th17细胞作为一种具有炎性效应的免疫细胞,特征性表达转录因子维甲酸相关核孤儿受体(retinoid-related orphan receptor gammar,RORγt),通过分泌IL-17等促炎性细胞因子在自身免疫性疾病、肿瘤以及感染性疾病中发挥重要作用[23]。研究发现在人和动物的慢性根尖周病损组织中均有Th17细胞相关细胞因子IL-17的表达[24-26],并且在根尖周病损组织中,与非活动性病损组织相比,活动性病损组织中IL-17的表达显著增高[27]。Yang等[28]利用实验性大鼠根尖炎模型,通过动态研究发现在根尖周组织的破坏过程中IL-17+细胞显著增多。以上研究结果表明IL-17在慢性根尖周炎的组织破坏中发挥重要作用。有研究发现在实验性小鼠系统性红斑狼疮模型中部分Treg细胞能够共表达转录因子Foxp3和RORγt,在微环境作用下转变成Foxp3+IL-17+T细胞,分泌IL-17,参与系统性红斑狼疮的进展和组织损伤。特异性阻断内源性Foxp3+RORγt+Treg细胞中RORγt的活化能有效地缓解病情和组织破坏[29-30]。Okui等[31]采用免疫荧光的方法在牙周炎的病损牙龈组织中检测到Foxp3+IL-17+细胞,并且通过流式细胞分析发现在病损组织中Foxp3+IL-17+细胞在Foxp3+细胞中的比例明显增高。上述研究结果提示Treg细胞具有可塑性,除具有免疫调节作用外,在一定条件下还可发挥促炎性效应。该研究中发现,虽然在慢性根尖周炎中Treg相关因子表达增高,但是不能有效地抑制根尖周组织的炎症进展和组织损伤,分析可能的原因有2个方面:一方面,Treg分泌的抑炎性细胞因子的效能不足,不能有效地对抗炎性细胞因子对根尖周组织的破坏作用;另一方面,Treg细胞在微环境作用下通过生成促炎性细胞因子,发挥促炎性作用,进而加重根尖周组织的炎症反应和组织破坏。
综上所述,在健康牙龈组织和慢性根尖周炎病损组织中均存在Treg细胞相关细胞因子FoxP3和IL-10的表达,并且慢性根尖周炎病损组织中Treg细胞相关细胞因子的表达显著高于健康牙龈组织,表明在慢性根尖周炎中Treg细胞介导的免疫反应增强。此外,Treg细胞分泌的抗炎性细胞因子的效能不足和Treg细胞产生的促炎性细胞因子的炎性作用可能是导致Treg细胞不能发挥有效抗炎作用的原因。推测Treg细胞在慢性根尖周炎的发病过程中可能发挥抑炎和促炎双向调节作用,但具体的作用机制还有待于进一步的研究。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
现代临床医学(2021年4期)2021-07-31
中国生殖健康(2019年6期)2019-01-06
知识就是力量(2018年6期)2018-06-15
小学生导刊(2018年4期)2018-04-18
中华老年口腔医学杂志(2016年2期)2017-01-15
中西医结合心脑血管病杂志(2016年20期)2016-03-01
中外医疗(2015年11期)2016-01-04
中国医疗美容(2015年1期)2015-07-12
医学研究杂志(2015年12期)2015-06-10
