
四川省猪源盖塔病毒的分离鉴定及遗传进化分析
2019-10-15 10:41江朝源曾喻兵彭珂楠张儒博杜佩珊殷鑫欢徐志文
中国人兽共患病学报 2019年9期
江朝源,李 飞,曾喻兵,彭珂楠,张儒博,杜佩珊,冯 宇,殷鑫欢,朱 玲,2,徐志文,2
盖塔病毒(Getah virus)是披膜病毒科(Togaviridae)甲病毒属(Alphavirus)成员[1]。1955年Scherer等[2]首次在马来西亚橡胶林中生存的白雪库蚊(Culex gelidus)中分离得到该病毒,GETV可感染多种动物并引起马和猪发病,其症状表现为发烧、荨麻样皮疹和后肢水肿[1],2017年10月周峰等[3]首次报道中国河南省某猪场感染盖塔病毒。该病毒粒子呈球形,有囊膜及纤突,直径约70 nm,单股正链RNA,病毒基因组全长约11~12 kp,编码区包含2个独立开放阅读框ORF1和ORF2;ORF1长约7 407 bp,编码4种非结构蛋白NSP1、NSP2、NSP3和NSP4;ORF2长约3 759 bp,编码5个结构蛋白Cap、E3、E2、6K和E1。5′端含有非编码区(5′UTR)和帽子结构,3′端含有非编码区(3′UTR)和Poly(A)尾[3]。由于衣壳蛋白Cap基因全长804 bp,该基因高度保守、对病毒颗粒的形成和存活至关重要[4];NSP3基因全长约1 572 bp,变异频繁[5],其C端参与病毒的复制[6],因此这2个基因常被分别用作该病毒的检测、鉴定和变异分析。
本研究首次从中国四川省一猪场混合感染猪繁殖与呼吸综合征(PRRS),临床症状表现为震颤、皮肤发红、精神颓废、棕黄色腹泻、关节炎的仔猪血液中检测并分离到1株GETV,对该病毒进行全基因序列测定,分析该病毒分子生物学遗传特征,对猪源GETV的感染谱和致病性、感染和传播机制、环境微生物学及公共卫生等相关研究具有重要意义,并为之提供宝贵的研究素材。
1 材料与方法
1.1 材 料
1.1.1 病料及细胞系 病料来源于四川省某发病猪场送检的发病仔猪血液,BHK21细胞、PK-15细胞由本实验室保存。
1.1.2 主要设备和试剂 RNAiso Plus来自宝日医生物技术(北京)有限公司;DL2000 DNA Marker、反转录酶试剂盒、2×TaqPCR Master-Mix、Nuceanse-Free water和p MD19-Tsimple载体,来自大连宝生物工程有限公司;Trizol LSReagent、小量质粒DNA提取试剂盒和小量胶回收试剂盒,于北京博迈德生物技术有限公司购买。大肠杆菌DH5α由四川农业大学动物生物技术中心制备,DMEM为Gibco公司产品、胎牛血清为北京四季青公司产品。PCR仪、Gel DocTMEZ imager凝胶成像系统购自Bio-Rad公司,恒温水浴锅、恒温摇床购自Thermo Fisher公司;甲基纤维素、福尔马林和结晶紫来自成都市科隆化学品有限公司。
1.1.3 检测引物的设计与合成 参考GenBank上GETV M1株全基因[7](登录号为:EU015061),生物工程(上海)公司利用Primer Premiers 5.0软件针对Cap和NSP3部分序列设计2对特异性引物,见表1。

表1 扩增GETV Cap和NSP3基因部分序列的引物信息Tab.1 Primers for amplification of some sequences of Cap and NSP3 genes of GETV
1.2 方 法
1.2.1 病料样品处理 取感染仔猪血样1 mL,用含青、链霉素的DMEM作3∶1(V/W)稀释。反复冻融3次后,3 000 r/min离心10 min,取上清保存到-80℃。
1.2.2 病毒分离鉴定 上清液经0.22μm滤器除菌,取0.2 mL接种到生长状况良好的单层BHK21细胞上。37°C吸附1 h,弃上清,加入2 mL含2%FBS的DMEM维持液,37℃、5%CO2条件下培养。每天观察记录细胞病变(CPE),若有病变收取病毒培养物,于-80℃反复冻融3次,离心取上清接种BHK21细胞进行传代培养。盲传3代后进行蚀斑纯化和RT-PCR检测。将纯化后有CPE且RT-PCR检测为GETV阳性的培养物用BHK21细胞、PK15细胞传代5次,收获病毒置-70℃保存;无CPE者丢弃。
将BHK21细胞(约106个/mL)接入96孔板长成单层后,弃去培养基,用无菌PBS冲洗3次。将第5代细胞培养物10倍系列稀释,各选取10-3~10-8共6个稀释度,以每孔100μL接种BHK21细胞,每个稀释度8个重复,同时设置阴性和阳性对照。37℃、5%CO2吸附1 h后,每孔补加100μL含2%FBS的DMEM维持液,观察并记录出现CPE孔数。试验重复3次,运用Reed-Muench法计算TCID50。
1.2.3 GETV核酸检测 取300μL病毒液按照TrizolRNA提取试剂盒说明书提取病毒总RNA。反转录体系(10μL):dd H2O 4.5μL、5×Buffer 2.0 μL、反转录酶0.5μL、d NTP 0.5μL、随机引物0.5 μL、模板2.0μL。反应条件为:35℃5 s、85℃15 min,cDNA模板用于PCR扩增。PCR体系(10 μL):2×TaqPCRmaster Mix 5.0μL、cDNA 1.0 μL、上游引物0.5μL、下游引物0.5μL、dd H2O 3.0 μL。反应扩增条件:Cap基因95℃预变性5 min;95℃变性30 s,56℃退火30 s,72℃延伸30 s,共35个循环,72℃延伸7 min;NSP3基因95℃预变性5 min;95℃变性30 s,56℃退火35 s,72℃延伸40 s,共35个循环,72℃延伸7 min扩增产物经1%琼脂糖凝胶电泳并进行凝胶成像观察。
1.2.4 GETV全基因扩增与测序 全基因组序列参考HNNY-2株设计12对引物分段进行扩增,扩增的目的片段按DNA胶回收试剂盒说明书回收纯化产物,与p MD18-T载体连接,转化、筛选及PCR鉴定后,送至生工生物工程(上海)公司测序,12对引物序列见表2。
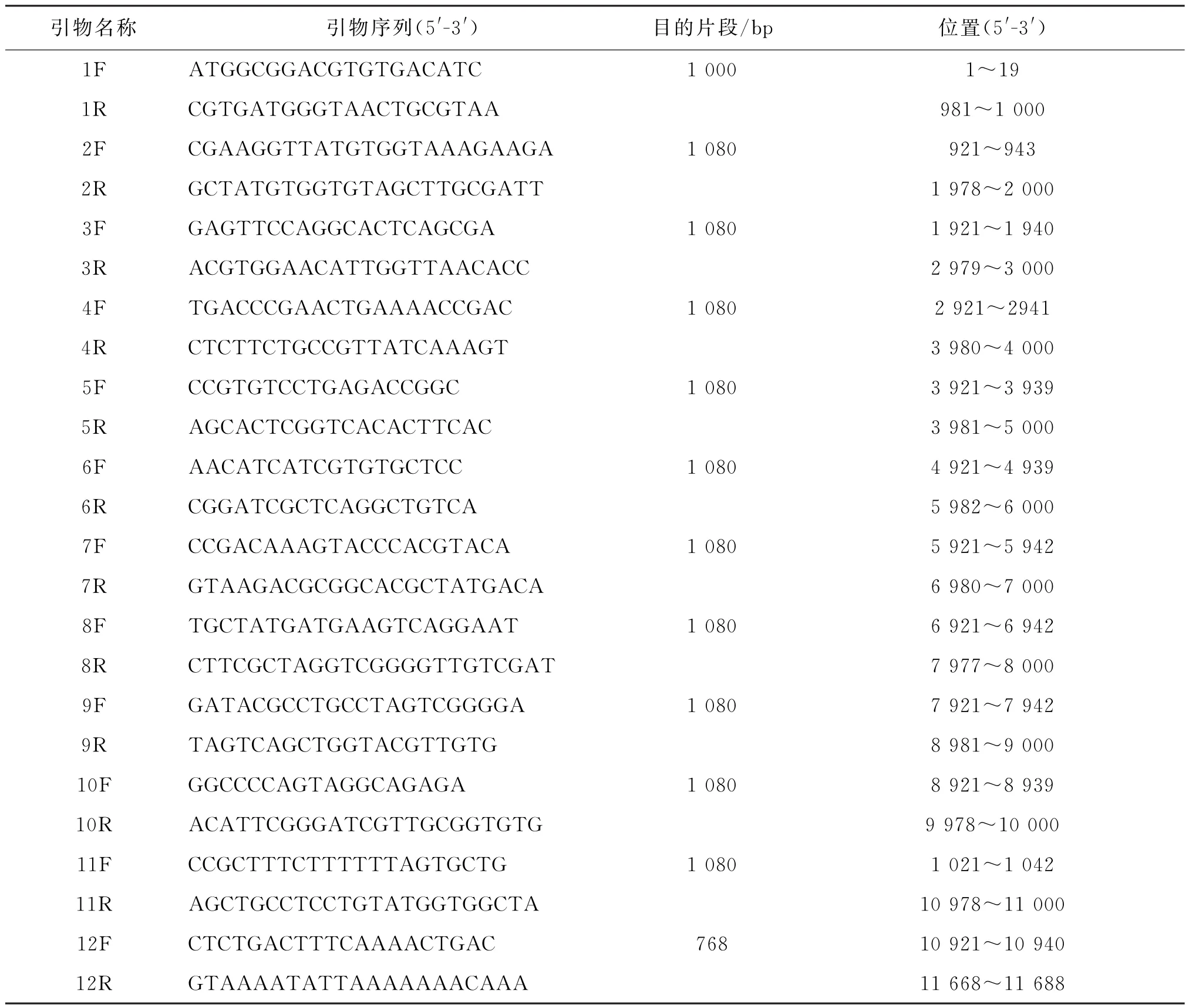
表2 扩增GETV全基因序列的引物信息Tab.2 Primers for amplification of all sequences of genes of GETV
1.2.5 GETV全基因序列分析 将测得的12个序列片段分别在NCBI数据库中进行BLAST比对,利用DNA Star将所有测序片段及5′和3′进行拼接,与GenBank上具有代表性的GETV毒株对序列进行相似性分析;用 MEGA 5.0中的邻接法进行遗传进化分析、绘制系统进化树。序列分析所用参考毒株见表3。
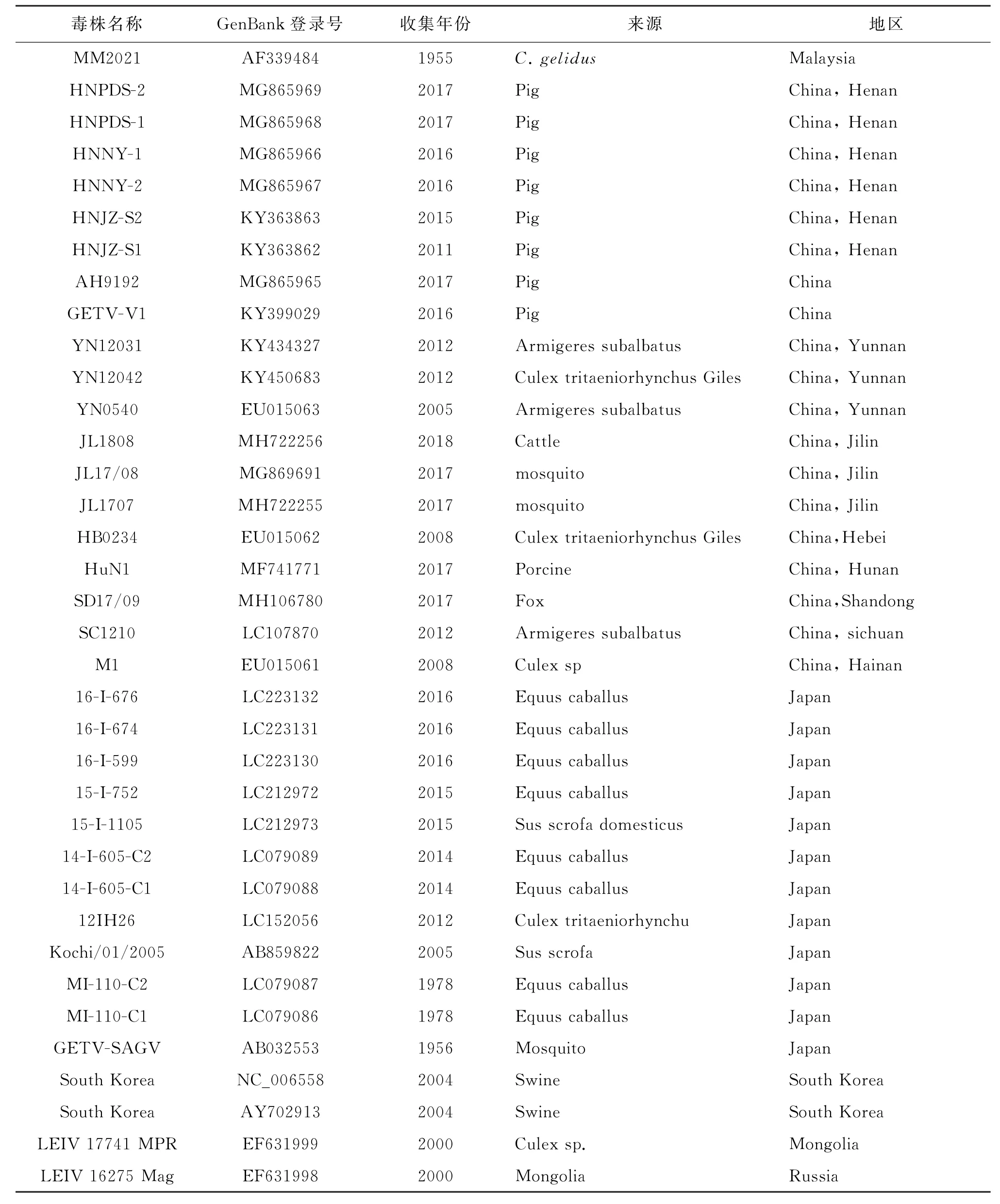
表3 国内外参考毒株Tab.3 Domestic and foreign reference strains
2 结 果
2.1 RT-PCR检测 病料样品、细胞培养物Cap、NSP3基因均为阳性。用GETV特异性Cap和NSP3基因引物对病料样品和第3代BHK21细胞培养物进行RT-PCR扩增,能够扩增出约316 bp、810 bp的预期单一目的片段(图1)。

图1 病料样品和第3代细胞培养物Cap、NSP基因RT-PCR产物扩增凝胶电泳图Fig.1 Gel-electrophoresis of disease material samples,Cap and NSP gene rt-pcr products of third-generation cell culture
2.2 分离到GETV 待测样品接种到BHK21细胞连续传代3次,经蚀斑纯化获得单个蚀斑克隆(图2)。同对照组相比,该分离物感染BHK21细胞和PK15细胞后约12 h,细胞出现空洞;24 h后细胞明显变圆,部分开始脱落;36 h后大量脱离,贴壁细胞大部分变圆(图3)。
细胞培养物经RT-PCR鉴定,可扩增出GETV的特异性目的条带,无PRRSV特异性目的条带。分离物传至第5代时用Reed-Muech氏法计算出细胞培养物的TCID50为106.85/0.1 mL将其分别命名为SC201807株。

图2 SC201807株感染BHK21细胞后的病毒蚀斑图Fig.2 Virus plaque of SC201807 strain infected with BHK21 cells
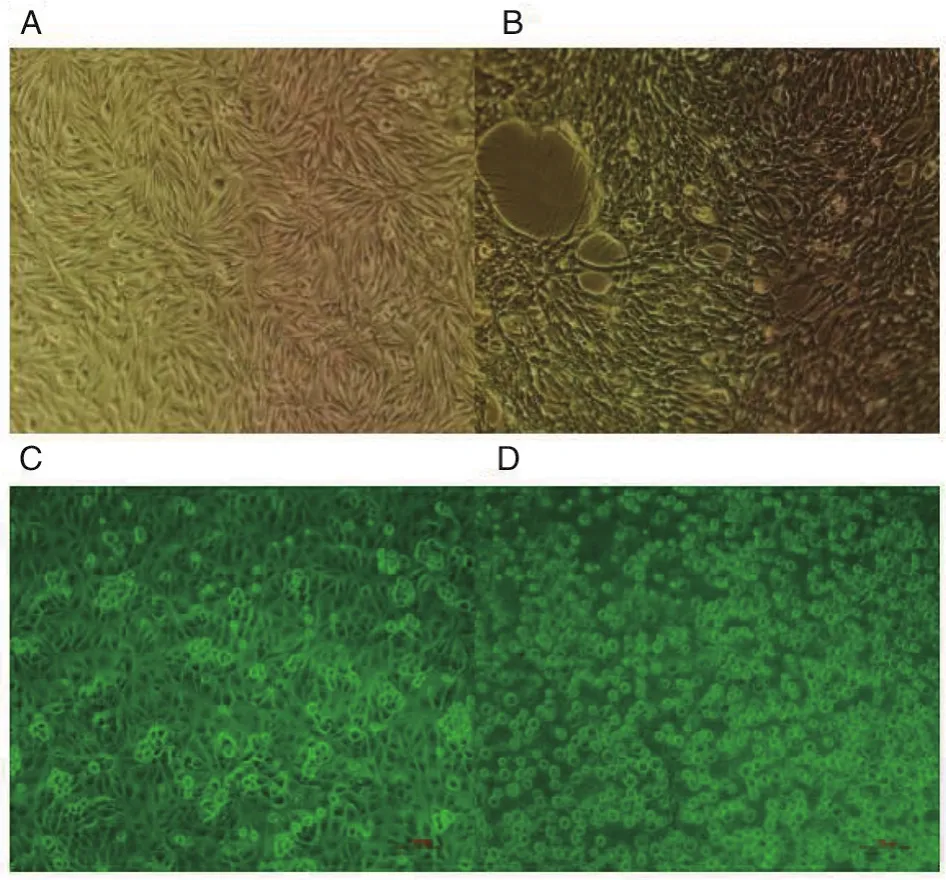
图3 SC201807株感染BHK21细胞、PK-15细胞后产生的细胞病变,放大倍数为10×20Fig.3 Cell changes after infection of BHK21 cells and PK-15 cells by SC201807 strain,the magnification is 10×20
2.3 获得SC201807毒株完整全基因组序列 利用12对特异性引物将SC201807基因组进行分段扩增,经回收、连接、转化、克隆、测序、拼接后,获得SC201801株全基因组序列,长度为11 691 bp。其碱基 T、C、A、G 占比例分别为19.7%、26.1%、28.0%和26.2%;GC含量为52.3%。基因组组成和开放性阅读框(ORF)大小与GenBank中GETV基本一致,基因组主要包括:5′UTR、3′UTR以及位于两者之间的两个开放阅读框,其中5′端起始的78个核苷酸和3′端最后403个核苷酸为非编码区、5′端非编码区之后的7 401个核苷酸编码构4种非结构蛋白(NSP1、NSP2、NSP3和NSP4),3′端非编码区之前的3 759个核苷酸编码5种结构蛋白(C、E3、E2、6K和E1),2个开放阅读框之间有44个核苷酸为非编码连接。SC201807株全基因序列已上传至GenBank登录号为MK693225。
2.4 GETV全基因组序列比对分析 对GETV全基因组核苷酸序列进行相似性分析,结果显示SC201807与GenBank中上传的35株GETV序列相似性为99.0%~96.1%,SC201807与中国河南省猪源毒株 HNPDS-2、HNPDS-1、HNNY-1、HNNY-2、HNJZ-S2、HNJZ-S1,中国吉林省蚊源毒株JL17/08、JL1707,河北省蚊源毒株HB0234以及日本蚊源毒株15-I-752、12IH26相似性最高为99.0%;与中国云南省蚊源毒株YN12031相似性最低为96.1%(图4)。

图4 SC201807株GETV与参考毒株全基因组核苷酸序列相似性(%)比较Fig.4 Comparison of nucleotide sequence similarity(%)between GETV strain and reference strain of SC201807
2.5 GETV E2蛋白氨基酸突变位点分析SC201807株E2序列的长度为1 266个核苷酸,在与36株GETV分离株E2蛋白氨基酸序列比对显示,SC201807株E2氨基酸序列与中国猪源HNPDS-2、HNPDS-1、HNNY-1、HNNY-2、HNJZ-S2、HNJZ-S1,中国蚊源HB0234以及日本蚊源16-I-676、16-I-674、16-I-599、15-I-752、15-I-1105、14-I-605-C2、14-I-605-C1、12IH26株高度保守,仅在E310(天冬氨酸→谷氨酸)发生突变。SC201807株与MM2021原始株比较共有4个氨基酸差异,分别为E54(苯丙氨酸→异亮氨酸),E73(络氨酸→组氨酸),E310(天冬氨酸→谷氨酸),E342(缬氨酸→异亮氨酸),见图5。

图5 SC201807株GETV与参考毒株E2蛋白氨基酸序列比对Fig.5 Comparison of amino acid sequences between GETV of SC201807 strain and E2 protein of reference strain
2.6 遗传进化分析 基于全基因组核苷酸序列遗传进化分析显示可以将GETV划分为4个主要进化群体,分别为1组M1株2008年中国海南分离所得,2组1956年日本分离到的GETV SAGV,3组2012年中国蚊虫中分离到的YN12031株,4组主要包括日本蚊源和中国猪源中分离出来的所有毒株,SC201807株也属于第4组,其与中国猪源HNJZS2及日本蚊源15-I-752、15-I-1105、14-I-605-C2、14-I-605-C1、12IH26亲缘关系最近,处于同一进化分支,如图6A。
通过E2推导的氨基酸序列遗传进化分析结果显示,可以根据E2大致划分为2个组,2012年中国云南分离到的YN12031株和2000年俄罗斯分离到的LEIV 16275 Mag株处于同一进化分支为1组;其余所有毒株为2组,SC201807属于2组与中国猪源HNJZ-S2及日本蚊源16-I-676、16-I-674、16-I-599、15-I-752、15-I-1105、14-I-605-C2、14-I-605-C1、12IH26亲缘关系最近属于同一分支,如图6B。
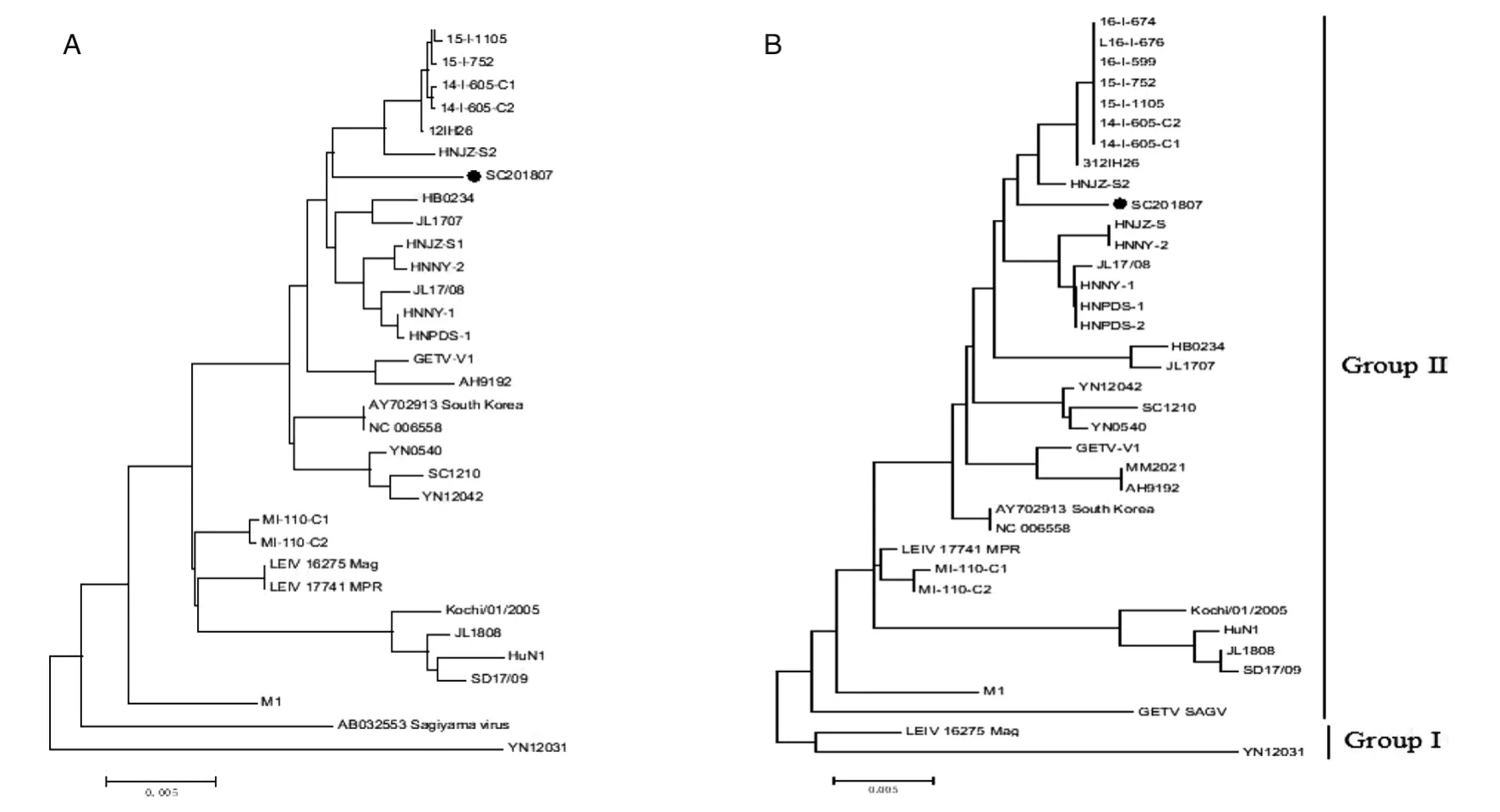
图6 SC201807株GETV与参考毒株全基因序列(A)及E2基因推导的氨基酸序列(B)进化分析Fig.6 Phylogenetic analysis based on the nucleotide sequences corresponding to the complete genome of isolate SC201807(A)and deduced amino acids sequences of E2gene(B)
3 讨 论
1955年发现GETV至今,血清学检测结果显示,GETV广泛分布在亚洲、大洋洲,在多种动物体内均能检测到GETV抗体,因此被确定为重要的动物源性病原体[8]。尽管尚未有引起人类疾病的报道,但人的血清样本中普遍能检测到GETV病毒中和抗体,因此目前人们一般把该病看做人兽共患病[9]。
目前我国对GETV的研究主要局限于人群中血清流行病学调查和库蚊体内病毒分离[10-12],至于蚊子体内的病毒从何而来以及动物在该病毒传播过程中所起的作用尚不得而知,GenBank中收录众多的中国来源GETV毒株,包括大量蚊源和个别猪源、狐狸源及蠓源[13]毒株。近两年来中国开始报道分离到猪源的GETV,由周峰等[3]于2017年报道分离到,并在山西、湖北、河南、安徽四省猪场中检测出37份阳性样本,同时2017年在湖南省猪群中发生了大规模GETV的感染,造成大量仔猪死亡,母猪流产并导致了重大经济损失[19]。
研究GETV的来源、在我国的生态学分布、对动物和人的致病性、确定其终末宿主以及蚊子是否在人和动物间扮演媒介角色等均具有重要的公共卫生意义和动物生产应用价值。本研究从四川某猪场流产胎儿体内分离到首株GETV,不仅扩大其感染谱,也为GETV可以感染猪群引起临床发病提供了充分证据,为今后深入开展其他相关研究提供了宝贵的研究素材,同时也为人群感染GETV的相关研究开辟了一片新领域。
我国作为世界养猪与猪肉消费第一大国,人们接触生猪及其产品的机会甚多,因此,猪病流行动态与人类健康密切相关。盖塔病毒属于人兽共患病,本研究不仅为公共卫生安全提出新的警示,也向探明该病毒的传播链条迈近了一步。另外值得注意的是,本研究是从混合感染猪繁殖与呼吸综合征的仔猪血液中分离得到的GETV,在进行分离鉴定时首先选用BHK21细胞进行分离并纯化,排除了PRRSV的干扰。在蒋正军猪繁殖和呼吸障碍综合征(PRRS)病毒囊膜糖蛋白(GP5)自身抗独特型抗体的鉴定报道中指出,感染PRRSV后1~2月才出现中和抗体,这就导致感染PRRSV后一定时期内免疫缺陷[20],PRRSV是一种免疫缺陷病,感染后机体免疫系统损伤,会造成混合或继发感染[14],推测本研究中的GETV和PRRSV混合感染可能是由于其免疫缺陷导致;而引起母猪流产的病毒种类甚多,包括日本脑炎病毒(JEV)、伪狂犬病病毒(PRV)、PRRSV、猪圆环病毒2型(PCV2)、猪圆环病毒3型(PCV3)猪细小病毒(PPV)和CSFV等。近年来病猪混合感染的概率不断攀升,其中PRRSV和PCV均属于免疫缺陷病,袁婧等[15]在报道中指出,PCV2与多种病原混合感染的现象十分普遍。推测GETV与这2种病毒的混合感染概率较高,王新港等[21]通过建立PRRSV与GETV的双重RT-PCR对河南省136份临床疑似PRRSV样品进行检测,共检出PRRSV阳性样品52份(38.2%),GETV阳性样品15份(11.2%),结果也表明GETV与PRRSV混合感染比例较高;在周峰等[16]流行病学调查报道中也证实了此推测,虽然GETV与各个病原混合感染之间的机制尚不清楚,但在临床研究中开展流行病学调查和建立混合感染检测方法,并分析其流行病学特征十分有必要。
将SC201807与Gen Bank数据库中现有的36株GETV全基因组核苷酸序列同源性比较发现,序列与近期公布的中国猪源毒株HNPDS-2、HNPDS-1、HNNY-1、HNNY-2、HNJZ-S2、HNJZ-S1以及日本蚊源毒株15-I-752、12IH26高度相似,即为99.0%,但与中国云南省蚊源毒株YN12031相似性最低为96.1%。通过对E2蛋白氨基酸序列突变位点分析发现GETV的结构蛋白E2基因高度保守[17],E2编码的结构蛋白是病毒的抗原识别位点和中和位点,在吸附靶细胞、感染宿主以及诱发机体产生抗感染免疫中充当重要角色,SC201807株E2氨基酸位点与中国猪源HNPDS-2、HNPDS-1、HNNY-1、HNNY-2、HNJZ-S2、HNJZ-S1,中国蚊源HB0234以及日本蚊源16-I-676、16-I-674、16-I-599、15-I-752、15-I-1105、14-I-605-C2、14-I-605-C1、12IH26株高度保守,仅在E310(天冬氨酸→谷氨酸)发生突变,氨基酸的稳定性更有利于基于E2蛋白开发的疫苗上产生交叉保护性。通过对全基因组核苷酸序列和E2蛋白氨基酸序列绘制遗传进化分析结果显示,SC201807株与中国猪源HNJZ-S2及日本蚊源15-I-752、15-I-1105、14-I-605-C2、14-I-605-C1、12IH26亲缘关系最近,处于同一进化分支;在李媛媛[17]的报道中将GETV大致划分为4个主要进化群体,除原始株 MM2021、GETV SAGV、YN12031其余所有从中国,日本,韩国和蒙古在1964年至2014年从蚊子,猪,马和其他动物中分离出来的所有毒株划分为同一组群,根据本研究的遗传进化分析中SC2101807株也属于此组群,同时中国猪源与日本蚊源的核苷酸序列高度同源以及在遗传进化中也处于同一分支表明中国猪源盖塔的传入与日本蚊源的相关性较大。
HiroshiBannai等[18]首次报道了2种不同动物马群与猪群之间盖他病毒感染与发病的地理空间和时间关系,表明2010-2015年期间猪群的盖他病毒阳性率在不断上升;而2014-2015年期间猪群与马群的GETV感染具有同步性;从猪群和马群分离的GETV其核苷酸全序列具有99.89%~99.94%的相似性。该研究结果提示上述2种家畜之间的GETV感染存在密切相关性,但究竟是什么因子在这种相关性中起媒介作用尚无确切证据。李媛媛等[22]对云南省家畜进行盖塔病毒血清学调查,发现其在鸡,鸭,奶牛,猪和肉牛的血清样本中均检测到GETV抗体,表明当地可能有大量的GETV宿主动物,媒介传播造成多种动物之间感染机制也尚不清楚。目前已知GETV有较强的宿主性,侵害多种动物的胎儿和新生儿,临床中重要的是监测GETV感染发生率,但因此如果只对正常人群进行血清学调查,而忽视对孕妇、产妇、新生儿及流产胎儿等重点对象进行深入研究,可能很难掌握该病毒对人类的真实危害程度。
尽管GETV进入人们视野已长达60余年,但对其危害及影响程度尚未清楚,随着各种相关研究的不断深入,相信人们将会对GETV具有更全面的了解和认识,从而为防治提出科学依据。
利益冲突:无
引用本文格式:江朝源,李飞,曾喻兵,等.四川省猪源盖塔病毒的分离鉴定及遗传进化分析[J].中国人兽共患病学报,2019,35(9):805-814.DOI:10.3969/j.issn.1002-2694.2019.00.112
猜你喜欢
动物医学进展(2022年9期)2022-11-26
军事文摘(2022年16期)2022-08-24
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
