
马梨形虫病病原双重荧光PCR快速检测方法的建立与应用
2019-10-12 01:23高志强赵相鹏
中国动物检疫 2019年10期
高志强,汪 琳,尹 羿,张 伟,蒲 静,赵相鹏,任 彤
(北京海关技术中心,北京 101113)
马 梨 形 虫 病(equinE piroplasmosis,EP)又称马焦虫病,是由蜱传播的马科动物血液原虫病。该病病原有两种,分别为马泰勒虫(Theileria equi)和驽巴贝斯虫(Babesia caballi)[1],其中的任何一种都可引起马科动物的急性感染,主要表现为发热、溶血性贫血、血红蛋白尿、黄疸、脾肿大,甚至死亡。有证据表明,马泰勒虫可经胎盘由母马传播给胎儿,造成流产、死产或新生幼驹梨形虫病[2]。
无明显临床症状的隐性带虫马科动物进入本病非疫区是造成本病传播的主要原因[2]。因此,各国想要保持无EP 状态,必须制定法规,防止感染动物进入。在国际马匹交易及相关活动中,均要求进行EP 检测[3-4]。对于隐性带虫马科动物,采用特异敏感的检测方法进行检测非常必要。
cELISA 方法目前已广泛应用于进出口马匹的检疫。马泰勒虫cELISA 检测方法,是使用了重组马泰勒虫裂殖子抗原1(EMA-1)蛋白和特异性单克隆抗体建立的,是检测动物感染抗体的成熟和敏感方法,能够检出不同地域的马泰勒虫抗体[5]。驽巴贝斯虫cELISA 检测方法,目前是使用了棒状体相关蛋白1(RAP-1)及其特异性单克隆抗体建立的[6]。
随着EP 病原分子生物学的发展,一些针对马泰勒虫和驽巴贝斯虫基因组特异位点扩增的检测方法也相继出现,如PCR、套式PCR、LAMP 以及定量PCR 等。本研究通过对国内外不同地理区域的马泰勒虫EMA1 基因以及驽巴贝斯虫18S rRNA进行序列比对,设计2 套带有不同荧光素标记的MGB 引物探针,经体系优化,建立了可同时检测马泰勒虫和驽巴贝斯虫的双重荧光PCR检测方法,并应用该方法,对10 份进口马全血和20 份马血清进行了检测。
1 材料与方法
1.1 材料
1.1.1 病毒核酸及被检样品 西尼罗病毒(West Nile virus)NY-99、马动脉炎病毒CVL Bucyrus、马流感病毒A/Equine/Xibei/1/2007(H3N8)、东部马脑脊髓炎病毒ssp. North American variant、西部马脑脊髓炎病毒McMillan 等病原的cDNA 以及马疱疹病毒1型、马链球菌兽疫亚种(C55100)的核酸,均为本实验室保存;2018 年进口的20 份马血清及10 份马全血,由本实验室收集保存。
1.1.2 主要试剂 血液/组织/细胞基因组提取试剂盒(DP304),购自天根生化科技有限公司;Ex HS Taq DNA 聚合酶、dNTP 等,购自TaKaRa 公司;核酸纯化柱及套管,购自上海生工。
1.1.3 主要设备 7500 荧光PCR 仪以及AB 9700 PCR 仪,均为AB 公司产品;毛细管电泳仪QIAxcel,为QIAGEN 公司产品。
1.2 方法
1.2.1 引物和探针合成 对于马泰勒虫,随机选取不同地域的EMA1 基因进行序列比对,选取保守位置,设计引物探针(图1);对于驽巴贝斯虫,则比对马泰勒虫和驽巴贝斯虫的18S rRNA 序列,仅在驽巴贝斯虫间保守的位置,设计引物探针(图2)。同时合成一对可用于扩增马泰勒虫的完整EMA1 基因引物,引物探针名称、序列及靶基因见表1。
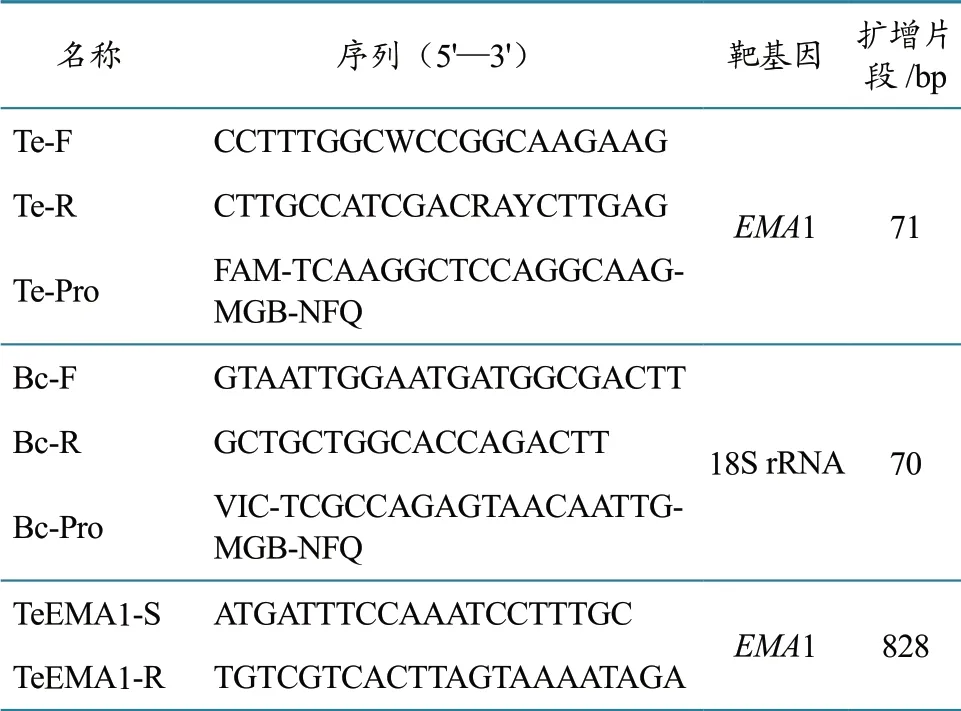
表1 引物、探针的名称、序列及靶基因
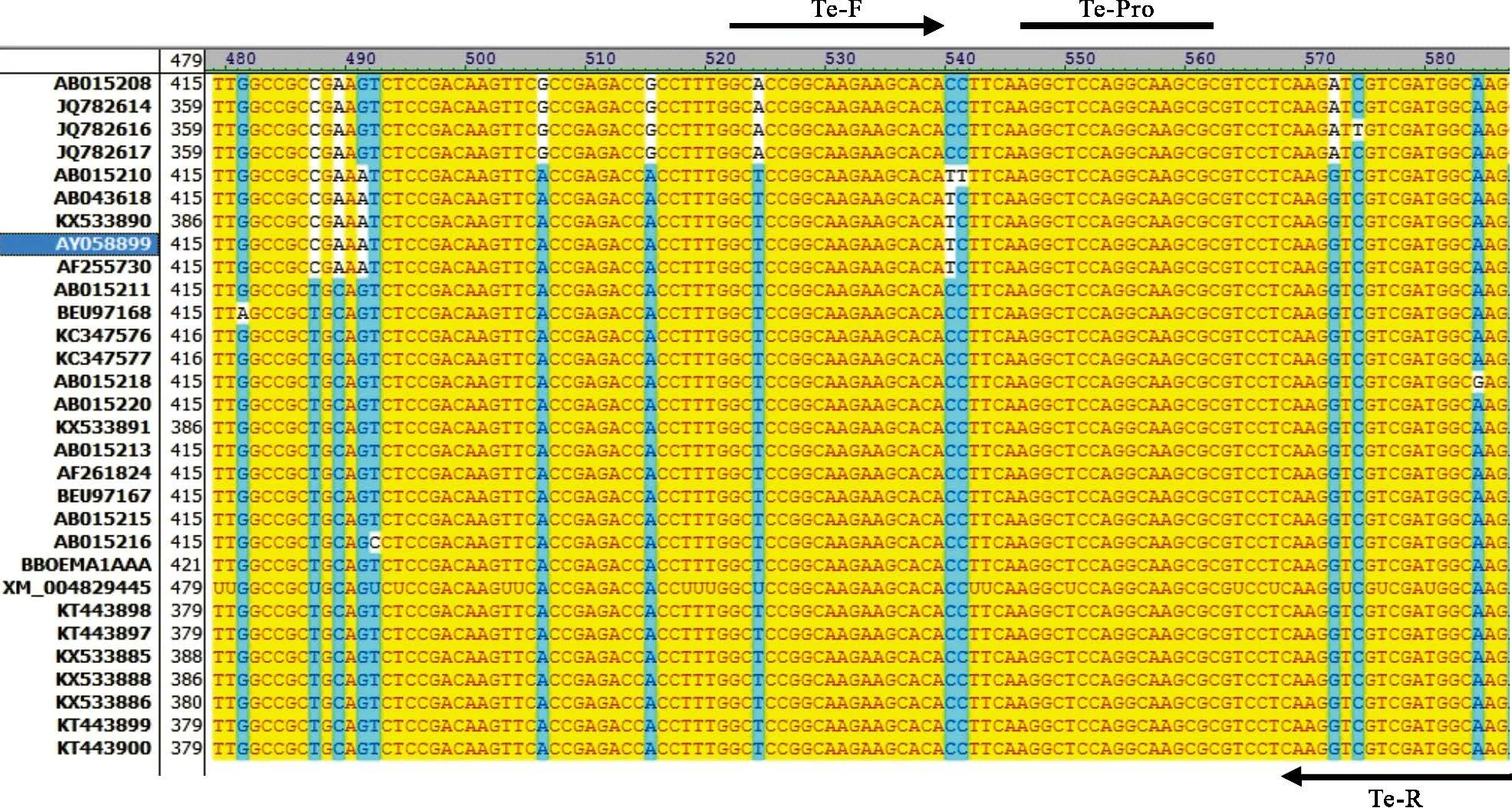
图1 马泰勒虫引物探针设计区域的序列比对

图2 驽巴贝斯虫引物探针设计区域的序列比对
1.2.2 反应条件优化 通过AB7500 设备,对各组分浓度进行优化,并对最优反应参数进行筛选。
1.2.3 灵敏度试验 将经测定计算的已知拷贝数的质粒DNA pGEM-T-EMA1 和pSKII-18SrRNA 作10 倍系列稀释,应用优化的反应体系进行检测,确定建立方法的分析灵敏度。
1.2.4 检测标准曲线建立 对质量浓度分别为8.7×10-4~8.7×103ng/mL 和1.0×10-3~1.0×103ng/mL的pGEM-T-EMA1 和pSKII-18SrRNA 的质粒DNA进行检测,以Ct 平均值为纵轴,质粒质量浓度为横轴,进行线性回归分析并作标准曲线,计算回归系数(R2)和PCR 扩增效率。
1.2.5 特异性试验 应用建立的反应体系,对西尼罗病毒NY-99、马动脉炎病毒CVL Bucyrus、马流感病毒A/Equine/Xibei/1/2007(H3N8)、东部马脑脊髓炎病毒ssp. North American variant、西部马脑脊髓炎病毒McMillan 等病原的cDNA 以及马疱疹病毒1 型、马链球菌兽疫亚种(C55100)的核酸进行检测,以验证方法的特异性。
1.2.6 样品检测 应用建立的方法,对20 份进口马血清及10 份马全血,进行实际应用检测。
1.2.7 马泰勒虫阳性样品EMA1 全基因扩增 使用表1 中的引物,对TeEMA1-S/TeEMA1-R,进一步对荧光PCR 检测为阳性的2 份马泰勒虫样品进行扩增,采用50 μL 体系,每个反应体系均包含1×PCR Buffer、3.0 mmol/L MgCl2、200 nmol/L dNTP、0.8 μmol/L TeEMA1-S/TeEMA1-R 引物以及2.5 U Taq DNA 聚合酶、5 μL 模板DNA。反应参数为:94 ℃ 3 min;94 ℃ 30 s,58 ℃ 30 s,72 ℃1 min,35 个循环;72 ℃最后延伸10 min。扩增后取PCR 产物5 μL,用2%琼脂糖凝胶电泳。
1.2.8 目的片段克隆、测序和序列分析 将PCR 产物经琼脂糖凝胶电泳后切胶回收目的片段,按照说明书克隆入pGEM-T 载体,转化Top10 感受态细胞,对每个阳性质粒各挑取2 个克隆进行序列测定。对EMA1 基因序列和推导氨基酸序列进行变异分析。将测定获得的2 个EMA1 基因序列与NCBI 获取的全部23 个马泰勒虫完整EMA1 基因序列进行比对,使用Lasergene 7.0 绘制进化树。
2 结果与分析
2.1 荧光定量PCR 反应条件优化
经优化确立的反应体系为:每个反应体系均包 含1×Ex TaqPCR Buffer、200 nmol/L dNTP、0.4 μmol/L 引物、0.2 μmol/L 检测探针。反应参数为:95 ℃ 3 min;94 ℃ 10 s,57 ℃ 30 s,72 ℃ 30 s,40 个循环。每个循环72 ℃时收集荧光信号。
2.2 灵敏度试验
该建立的双重反应体系可以检出22 拷贝的马泰勒虫质粒DNA(图3-A)和31 拷贝的驽巴贝斯虫质粒DNA(图3-B),表明方法的灵敏度可以满足要求。
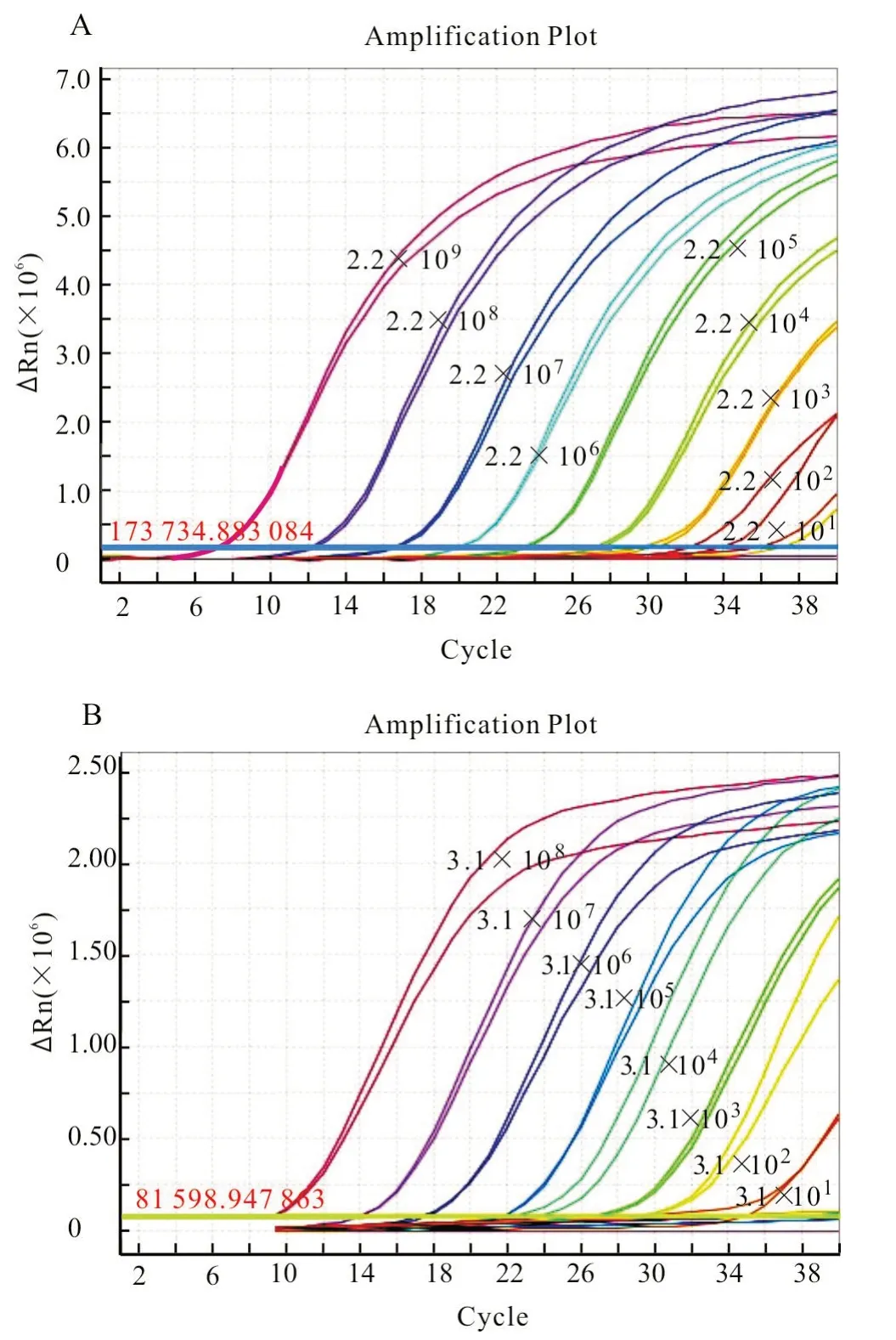
图3 灵敏度试验结果
2.3 回归与荧光PCR 效率计算
对已知浓度的质粒DNA 进行检测,将检测结果经计算后进行线性回归,获得的相关数据及方程图形见图4,发现PCR 反应效率分别为92.125%和98.088%,表明建立的方法扩增效率较高。
2.4 特异性试验
应用建立的检测反应体系,对西尼罗病毒NY-99、马动脉炎病毒CVL Bucyrus、马流感病毒A/Equine/Xibei/1/2007(H3N8)、东部马脑脊髓炎病毒ssp. North American variant、西部马脑脊髓炎病毒McMillan 等病原的cDNA 以及马疱疹病毒1 型、马链球菌兽疫亚种(C55100)的核酸进行检测,结果显示建立的检测方法与上述病原核酸无交叉反应。

图4 回归分析及获得的方程图形
2.5 送检样品检测
应用建立的方法,对2018 年进口的20 份马血清及10 份马全血进行检测,结果检出2 份马全血为马泰勒虫阳性(编号8 和9);进一步使用毛细管电泳,对PCR 产物进行分析,观察到了预期大小的目的片段(图5)。
2.6 阳性样品的完整EMA1 基因扩增与克隆
采用TeEMA1-S 和TeEMA1-R,经PCR 成功扩增了8 号和9 号全长 EMA1 基因(图6);将其克隆入pGEM-T,质粒经菌落PCR 鉴定后,证明2 个样品的EMA1 基因成功克隆入质粒载体,分别命名为pGEM-T-EMA1-8 和pGEM-T-EMA1-9。
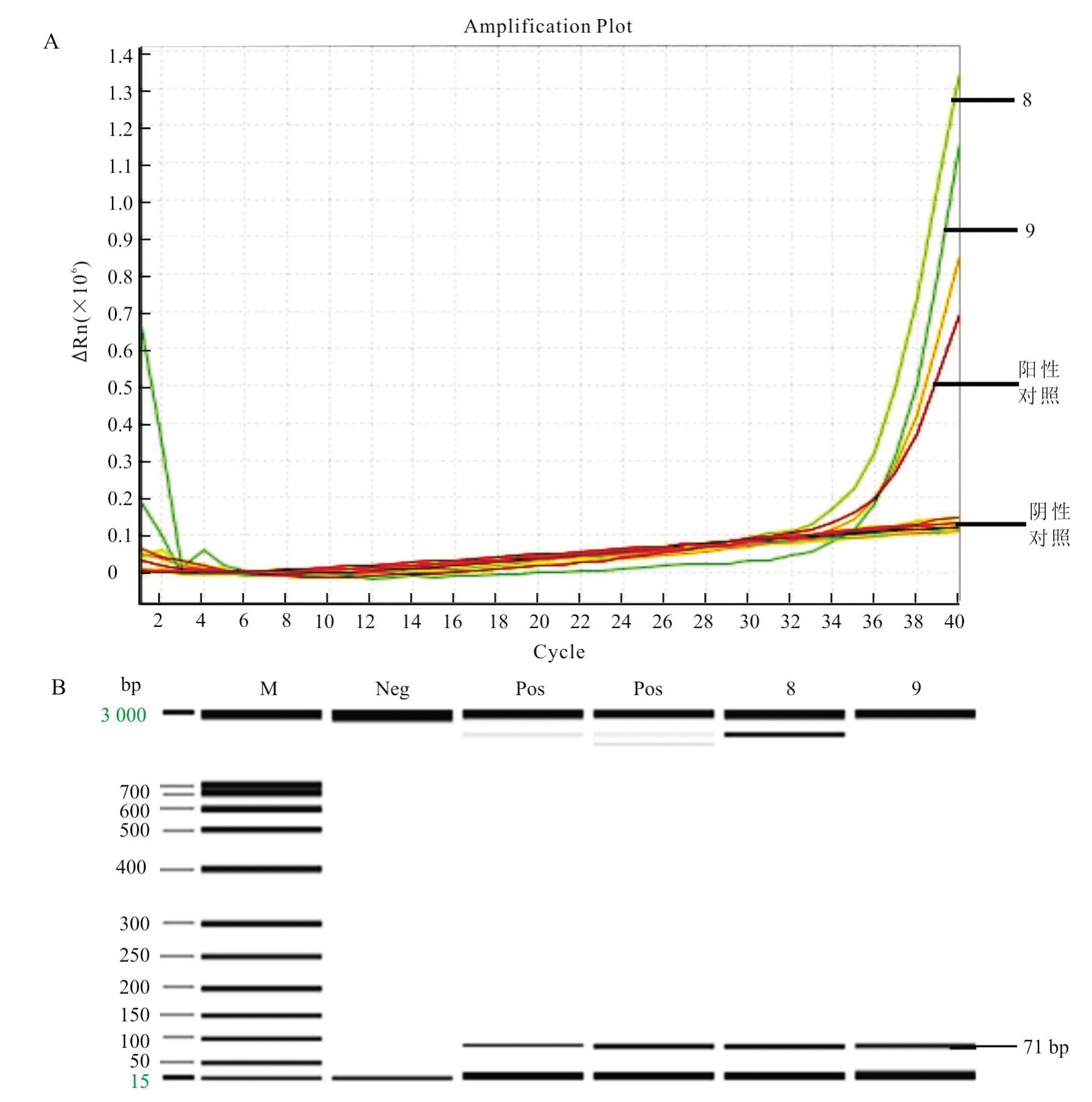
图5 送检样品的荧光PCR 检测结果

图6 2 个样品EMA1 基因PCR 扩增结果
2.7 序列测定与变异分析
测序结果经blast 分析显示,目的片段均为马泰勒虫EMA1 基因,与预期一致,长度均为819 nt,编码273 个氨基酸,二者序列相似性为98.9%。对其推导氨基酸序列进行比对,发现存在6 处氨基酸突变,显示两个样品中的EMA1 基因并不完全一致(图7)。
2.8 进化分析和亲缘性分析
使用Lasergene 7.0 MegAlign,将测得的2 个EMA1 序列与NCBI 获取的全部23 个马泰勒虫完整EMA1 基因序列进行了比对,并绘制了进化树(图8)。结果显示,2 个序列处于不同分支,提示基因可能来自不同的马泰勒虫虫株。

图7 EMA1 推导氨基酸序列比对结果
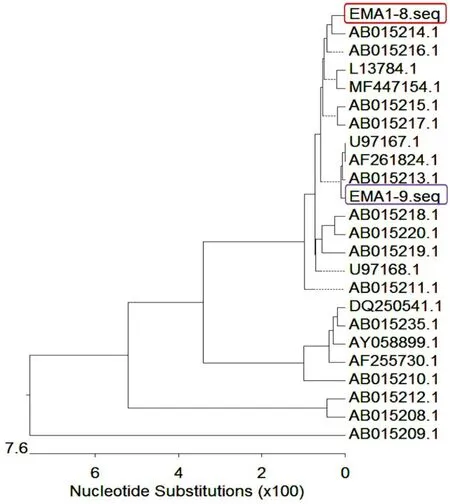
图8 EMA1 序列的进化分析结果
3 讨论
本研究针对马泰勒虫EMA1 基因和驽贝贝斯虫18S rRNA 靶基因,采用MGB 双标记探针,建立了一套可快速检测2 种马梨形虫病病原的双重荧光PCR 方法,并初步评价和验证了方法的有效性。研究显示,该方法灵敏、特异,与其他马病病原核酸无交叉反应。应用建立的方法,从2 份马泰勒虫抗体阳性全血中检出虫体核酸;进一步对2 份阳性样品的马泰勒虫EMA1 全基因进行扩增、克隆和序列分析,发现2 个序列存在差异,表明其可能来自不同的马泰勒虫虫株。
有研究显示,马泰勒虫流行地区的cELISA 确认阳性病例往往虫体核酸检测也呈阳性,而且抗体阴性马匹也可能呈虫体核酸阳性,表明核酸检测方法对马泰勒虫具有较高的诊断敏感性,而且核酸检测可能比cELISA 方法具有更高的敏感性[7]。本研究对2 份马泰勒虫cELISA 抗体阳性动物的全血检测为虫体核酸阳性,与相关报道一致,显示马血中马泰勒虫抗体和虫体可同时存在。研究还表明,目前马泰勒虫核酸检测的靶基因主要为18S rRNA 和EMA1 基因,二者检测结果基本一致。而马泰勒虫EMA1 特定序列在虫体内为单拷贝,因此本研究选择了该基因进行引物探针设计。由于建立方法的灵敏度和PCR 反应效率较高,因此针对该基因的荧光定量PCR 方法可用于虫体的定量分析。
对于驽巴贝斯虫,许多研究表明,cELISA 和核酸分子检测方法的一致性较差。巴西的一项调查显示,抗体阳性马匹中,18S rRNA 定量PCR 检测的阳性率仅为13.5%[7]。核酸阳性率与抗体阳性率差异较大也见于水牛的牛巴贝斯虫(B. bovis)和双芽巴贝斯(B. bigemina)感染[8]。目前对造成这种检测差异的原因还没有彻底弄清。一些研究显示,驽巴贝斯虫感染与马泰勒虫不同,宿主可能会自行清除驽巴贝斯虫虫体,但持续性的抗体检出可能是其他虫体感染引起的非特异性反应。本研究对2 份驽巴贝斯虫抗体阳性马的核酸检测结果为阴性,原因不明。
由于马梨形虫病病原感染的靶细胞为红细胞,因此血清中一般不能检出病原核酸,因此只能采集抗凝血进行检测。但由于一些单位采用肝素抗凝管采集全血,而肝素对PCR 的抑制作用较强,而且一般的核酸提取方法并不能去除肝素,因此检测结果往往为阴性。本研究同样发现血清的核酸检测结果均为阴性,而且发现肝素对PCR 的抑制作用较强,但通过洗涤血细胞和聚乙二醇沉淀法处理提取DNA,可显著消除肝素对PCR 的抑制作用。
猜你喜欢
贵州畜牧兽医(2022年1期)2022-03-07
临床肺科杂志(2021年5期)2021-04-27
数理报(学习实践)(2021年5期)2021-04-07
饲料博览(2020年6期)2020-12-30
北京航空航天大学学报(2017年2期)2017-11-24
发明与创新·中学生(2016年8期)2016-05-14
卷宗(2014年7期)2014-08-27
世界文学评论(2014年2期)2014-04-12
英语学习·新锐空间(2013年1期)2013-05-08
环球时报(2009-07-20)2009-07-20
