
甾体激素药物的生物转化研究进展
2019-10-10 02:29徐慧静崔立迁牛建娜
生物加工过程 2019年5期
徐慧静,刘 萍,崔立迁,牛建娜
(1.天津市食品安全检测技术研究院,天津 300308;2.天津市产品质量监督检测技术研究院,天津 300384)

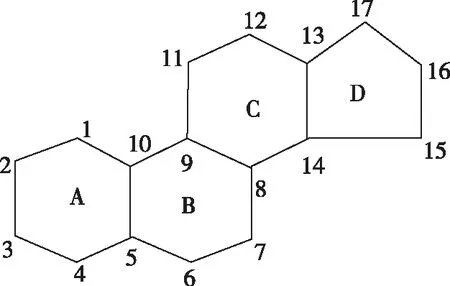
图1 环戊烷多氢菲核Fig.1 Cyclopentane polyhydrophenanthrene
甾体化合物被誉为“生命的钥匙”,可调节人体糖代谢、脂肪代谢、蛋白质合成、矿物质代谢及性功能[4]。甾体化合物具有分子量小、脂溶性大的特点,使其易于进入细胞并产生一系列生理变化,因此,甾体化合物具有成为药物的自然属性[5]。在临床上,肾上腺皮质激素是治疗类风湿性关节炎、支气管哮喘、结缔组织病变和湿疹等疾病的首选药物[6]。此外,甾体激素还具有保健的生理功能,如改善蛋白质代谢、抗肥胖、恢复和增强体力、预防冠心病及利尿降压等[7]。近年来,甾体药物的应用范围快速扩大,在治疗细菌性脑炎、淋巴母细胞白血病、人体器官移植、预防艾滋病毒(HIV)的传染和治疗内分泌疾病等方面也有相应用途[8]。甾体激素类药物现已广泛应用成为临床上仅次于抗生素的第二大类药物。
随着人们对甾体药物的不断研究,甾体药物的高效合成具有非常重要的价值和意义。与化学合成相比,甾体药物的生物转化具有如下的特点:①步骤少,周期短;②收率高,副产物少;③专一化强,快速精确地实现化学法难完成的反应;④立体选择性和区域选择性好;⑤绿色安全,无环境污染,条件温和。目前生物合成已经成为工业上合成甾体药物的重要途径。在甾体药物的生物转化生产过程中,微生物几乎能在甾体化合物的各个位点发生作用,包括甾体母核、C18和C19的两个角甲基及其侧链(图2)。

图2 甾体化合物的主要生物转化位点Fig.2 Major sites of biotransformation reactions for steroids
植物甾醇是我国类固醇药物合成的主要原料,且来源广泛。精炼油脂废渣可作为工业化生产类固醇药物的原料,既可提供充分来源又可变废为宝,减轻环境污染。甾体化合物的微生物转化反应类型[9]如表1所示。甾醇的微生物降解过程中存在三方面的问题:①甾醇疏水性强,分散性差,在水溶液中溶解度低;②甾醇核的降解,产物的进一步降解以及作为微生物利用碳源的甾醇核B环的自发破坏作用;③产物对细胞的毒性作用。如果甾醇以粉末投料会影响甾醇的利用率,且甾醇微生物降解酶是胞内酶,所以在甾醇使用前,需要将甾醇通过相应的媒介扩散到细胞中方能被利用。因此,提高甾体转化率的研究主要集中在两个方面:一是使用细胞固定化技术或不同的反应过程来改善类甾体底物的水溶性,提高水相中甾体底物的传质效率;二是利用现代生物技术转化用于生物转化的微生物,提高酶的活性,提高酶反应的选择性[10-11]。

表1 甾体化合物的微生物转化反应类型[9]
1 甾体化合物的微生物转化
1.1 微生物转化法的研究
皮质激素是第一个微生物转化的甾体药物,它的转化是生物转化的里程碑,开创了微生物转化甾体化合物的先河。早在1952年Peterson等利用Rhizopusnigricans中的羟化酶成功的生物转化黄体酮[12],在C11位上引入11α-OH,转化率高且专一性强,克服了皮质激素合成中难以引入11α-OH的困难,实现了化学方法难以实现的反应。Patrice等分别对可的松与泼尼松进行了3种真菌的生物转化[13],结果表明,氧孢镰刀菌UAMH9013中可能存在C7和C15羟化酶。而将这些甾体化合物作为ExophialajeanselmeUAMH 8783的底物,结果显示该真菌中存在高活性的5α和14α羟基化酶,同时具有氧化烯丙基的能力。CeratocystisparadoxaUAMH 8784对上述甾体化合物进行转化,结果显示单氧酶影响烯丙基的羟基化并且具有BV氧化重排的活性,然而,氧化还原反应起主导作用。Restaino等[14]利用玫瑰产色链霉菌(Streptomycesroseochromogenes)研究了氢化可的松的生物转化过程,在C16位引入16α-OH,得到16α-羟基氢化可的松,它是生产丙酸羟泼尼龙的重要中间体。此外,蓝色犁头霉(Absidiacoerulea)和新月弯孢霉(Curvularialunata)还可以对醋酸可的松和醋酸泼尼松的C11位进行羟基化,再次为皮质激素的生产提供了基础性的突破[15-17]。
甾体激素药物的关键中间体为雄烯二酮(AD)与雄二烯二酮(ADD),大部分甾体药物的生产都以这两种中间体为原料。目前对于AD的生产,主要通过植物甾醇的生物降解,如张亚楠等[18]开发了利用工业用菌株野生小金色分枝杆菌(Mycobacteriumneoaurum)催化降解甾醇制备4-AD的无油生产工艺,AD进一步脱氢即可获得ADD。大多数真菌对上述两种化合物具有甾基化催化活性,例如:Isariafarinosa具有广谱的高效类固醇羟化酶,所得到的7-羟基脱氢表雄酮具有较高的生物活性,可用于治疗阿尔茨海默病,是合成醛固酮拮抗剂的关键中间体[19]。该菌生物转化孕激素可得到6β,11α-dihydroxyprogesterone,是一种生理活性非常高的孕激素。Kozowska等[19]从波兰下西里西亚Voivodeship地区的废弃矿井中采集到了12株昆虫病原丝状真菌Isariafarinose,这12菌株均能有效转化脱氢表雄酮(DHEA),其中IsariafarinosaKCh KW1.1 可高效转化ADD、雄烯二酮、肾上腺甾酮、17α-methyltestosterone、17β-hydroxyandrost-1,4,6-triene-3-one和黄体酮。王雪荣[20]筛选出具有4-AD羟基化能力的4株丝状真菌:斜卧青霉(Penicilliumdecumbens)、曲霉菌(Aspergillussp.)、雷斯青霉(Penicilliumraistrickii)和赭曲霉(Aspergillusochraceus),但是这4株菌对4-AD的羟基化位点不同,所产生的产物的生理活性也有差异。张广求等[21]研究了刺盘孢属菌YNCA0116对孕酮和4-AD的生物转化,孕酮被该菌转化为11α,15α-二羟基孕酮和6β,11α-二羟基孕酮,4-AD被转化成11β,15α-二羟基-4-雄烯二酮,确定了刺盘孢菌YNCA0116具有转化甾体二羟基化的能力,但是对不同甾体化合物羟基化特点不同。
雷斯青霉(Penicilliumraistrickii)具有15α羟基羟化酶的特异性,可以催化左旋乙基甾烯双酮15α位羟基化生成15α羟基左旋乙基甾烯双酮,而且转化率可以达到93%以上,该工艺是合成新一代避孕药孕二烯酮所必需的。Torshabi等[22]利用Mucorracemosus生物转化甲基睾丸素(MT),最终可生成7α-OH-MT、15α-OH-MT和12,15α-OH-MT这3个产物。生物转化16α,17α-环氧黄体酮(EP)的研究过程,主要应用Rhizopusnigricans[23]、aspergillusochraceus[24]、Curvularialunata[25]、Cunningnamellaelegans[26]、Absdiacoerulea[27]和Metarhiziumsp.[28]等菌株催化EP在C11位的羟基化。此外,ColletotrichumliniAS3.4486还可以催化EP生成双羟基产物1β,15α-OH-EP[29-30]。但目前这些研究,还都处在实验室研究阶段,离工业化应用还有一段距离。
Janeczko等[31]以毛壳菌(Chaetomiumsp.1 KCH 6651)作为生物催化剂,黄体酮被高效转化为6β,14α-二羟基雄甾-4-烯-3,20-二酮,而脱氢表雄酮先被转化为7α-羟基衍生物,随后转换7-羟基雄甾-4烯-3,17-二酮。Gao等[32]利用丝状真菌Penicilliumcitrinum对3β-乙酸基孕甾-5,16-二烯-20-酮进行转化,生成4种代谢产物,分别为3β,7β-二羟基-孕甾-5,16-二烯-20-酮、3β-羟基-7α-甲氧基-孕甾-5,16-二烯-20-酮、3β,7β,11α-三羟基-孕甾-5,16-二烯-20-酮和3β,7α-二羟基-孕甾-5,16-二烯-20-酮,其中7-O-甲基化是孕甾生物转化的一个新反应。Kolet等[33]利用Mucorhiemalis生物转化黄体酮,得到14α-羟基黄体酮、6β,14α-二羟基黄体酮和7α和14α-二羟基黄体酮3种产物,总转化率达到94%,该研究一次反应得到3种羟基化黄体酮衍生物,并且这3种物质均具有工业开发潜力,是生物转化中比较理想的研究结果。
工业上重要的甾体化合物微生物转化反应类型总结见表2。
1.2 微生物代谢甾醇的分子机制
甾体降解是有氧代谢[34],早在20世纪八九十年代,研究人员就能够根据降解过程中产生的中间产物来研究出可能的代谢途径[35]。随着甾体降解基因簇[36]的发现,关键酶[37]的功能已被逐一确定。基因簇中的大多数基因与甾体代谢途径有关,其多步降解机制已被人们成功解释。

表2 工业上重要的甾体化合物微生物转化反应类型
通过对微生物代谢中间体的鉴定,构建了甾醇生物降解的可能代谢途径,并通过诱变去除了该途径中的关键酶,从而积累中间产物[38]。然而,人们逐渐发现,这种传统的方法会遇到菌株选育的瓶颈。由于诱变剂的成功率低、作用弱,无法解决突变体和代谢副产物的稳定性问题,很大程度上限制了这类微生物的应用。随着分子生物学的发展,人们试图从代谢工程和基因工程等多角度揭示红球菌[39]和分枝杆菌[40]等降解甾醇的分子机制,实现人工修饰和调控。对此,van der Geize团队的Rosåoniec等[41]揭示了红球菌和分枝杆菌甾醇降解的关键基因簇,并解释了一些关键基因的功能。同时为研究分枝杆菌感染性的致病机制提供了重要参考[42]。到目前为止,大多数典型的分枝杆菌和红球菌已经被测序,其中一些还通过甾醇诱导的转录组和基因芯片进行了检测[43]。杜大庆[44]研究发现,微生物体内与甾体代谢相关的基因具有集群化趋势聚集成簇,在甾体诱导下,大多数簇内基因均显著上调。在红球菌RHA1[45-46]中,有6个基因簇被表达上调,其中1个尤为显著。Petrusma等[39]和Kendall等[40]研究发现,该簇类包含51个基因,其中大部分与功能分析鉴定的关键基因高度同源,被鉴定为具有特异性的甾体代谢基因簇。
1.3 生物转化甾醇的方法
甾体化合物的微生物转化过程一般由两个阶段组成。第一阶段是对菌体进行培养,通过为菌体提供丰富的营养,使菌体能够在最适宜的温度、酸碱度和通风的条件下生长,以获得足够量的菌体和酶。培养时间的长短会因菌种的不同而不同,一般来讲,细菌通常需要 12~24 h,霉菌需要24~48 h。第二阶段是向培养液中加入甾体化合物来完成生物转化的过程。
1.3.1 全细胞转化法
全细胞生物催化在甾体化合物的工业生产中占有主要地位。具有甾体化合物催化能力的酶一般都是氧化还原酶类,通常位于膜蛋白上,在催化过程中还需要辅酶的参与。与酶催化相比,全细胞催化避免了成本昂贵、步骤繁琐的酶的纯化步骤,而且不需要在体系中额外添加昂贵的辅酶,因此大大降低了生产成本。
在自然界中,已发现许多微生物如细菌、真菌、藻类等可以对甾体化合物进行生物转化。简单节杆菌(Arthrobactersimplex)、分枝杆菌(Mycobacteria)、棒状杆菌(Corynebacteria)、假单孢杆菌(Pseudomonas)均可以对甾体母核C1,2位进行脱氢。尖孢镰刀菌(Oxysporum1301)菌株可催化多种甾体化合物,且生成睾内酯的产率较高[47]。在众多的微生物中,真菌对甾体药物的转化最为普遍,而且绝大部分都是进行羟基化反应,常用的菌种见表3。
1.3.2 静息细胞转化法
静息细胞转化是通过制备菌体静态悬浮液来进行的生物催化技术。将菌体培养到对数生长期后,过滤或离心收集菌体,再将菌体重悬于水中或适宜的缓冲溶液中,然后加入底物再进行生物转化反应。该方法具有如下优点:底物和菌体量的比例可变;杂质相对较少,易于分离纯化;反应时间短;菌体可多次重复利用等。例如,当诺卡氏菌氧化胆固醇时,湿细菌的酶活性可以保持在-20 ℃以下,并长期保持稳定。徐阳光[48]利用新金分枝杆菌生长细胞和静息细胞转化体系,对植物甾醇转化过程中产物母核降解问题进行研究,发现在静息细胞转化体系中,提高转化温度,甾醇能够正常转化,并且产物降解受到明显抑制,转化3 d时,产物降解率由38.31%下降为16.22%。通过改用AD作为底物,发现反应2 d后AD的降解率减少约50%。

表3 用于甾体化合物羟基化的微生物
1.3.3 固定化细胞转化法
固定化细胞技术既可以增加菌体浓度又可以重复利用菌体,既能提高催化效率又有利于生物催化剂与产物间的分离。固定化细胞技术在甾体生物转化上的应用实现了甾体药物的半连续化生产。菌体培养后,投入底物进行转化,随着转化时间的增加,菌体会发生自溶现象,而采取固定化细胞的方法可以解决该问题。固定化微生物细胞的方法很多,主要包括包埋及交联、载体吸附、共价结合等。
固定化细胞的在甾体生物催化中的效率主要取决于所用载体的性质如表面、大小、空隙度及亲水性等,另外反应条件、固定化方式也会对固定化细胞的活性产生重要影响。例如:将新月弯孢霉(Curvularialunata)先进行底物诱导,收集后的湿菌体固定在聚丙烯酰胺凝胶中,可重复多批次地催化 C11-β羟化反应[49]。而利用聚丙烯酰胺、海藻酸等固化剂等进行简单节杆菌的固定,能高效地进行甾体化合物的C1,2脱氢反应,且批次酶活稳定,能够长期使用。另外也可将固定化细胞装在玻璃柱中组成反应柱使反应液连续通过,有效进行C1,2脱氢反应。
2 甾醇侧链降解反应
早在1985年,Owen等[50]就对假单胞菌降解甾醇侧链产生AD的机制进行了研究,经测定,该工艺共分16步,涉及11种酶催化剂。到1990年,Szentirmai[35]进一步阐明了分枝杆菌NRRL B-3805对P-谷甾醇侧链降解产生AD的过程和机制,其过程主要包括C3羟基氧化脱氢、甾体核上C5双键异构化、C17烷基侧链降解。反应的顺序还有待确定,但可以肯定C17侧链的降解是其最关键的步骤[51]。
为了更清楚地了解11种功能酶参与甾醇侧链降解和酶反应的机制,Thomas等[52-53]、Wilbrink等[54]、Griffin等[55]和Yao等[56]研究过一些关键酶的基因和代谢机制。特别是Griffin等[55]通过高通量转座子突变发现了几个直接参与基因簇内外侧链降解的关键基因。Yao等[56]首次阐明了胆固醇氧化酶ChoMl和ChoM2在分枝杆菌ATCC 25795产生AD (D)过程中的代谢功能、分子机制及重要作用,并提高了关键酶ChoM2的表达,使AD产量增加51.2%。
然而迄今为止,甾体相关的研究基本上都是基于基因突变或敲除技术来定位甾醇侧链降解过程中的部分关键基因和酶,对全部过程中涉及的11种功能酶还没有全面的了解,目前甾醇的侧链降解只能在微生物的全细胞内进行,不能实现纯酶催化。
3 生物转化体系强化甾体生产效率
3.1 水相体系
甾体化合物在水中溶解度极低,极大地限制了底物传质进而影响底物与酶的接触,细胞壁和细胞膜作为细胞质的保护屏障,限制了底物进入胞浆的效率,也会降低生物转化的产率[57-58]。万古霉素、卵磷脂、氨基乙酸、鱼精蛋白、乙胺丁醇、多粘菌素B九肽、聚乙烯亚胺及杆菌肽等都可导致相关菌株细胞壁的缺失[59-61]。
3.2 双液相体系
双液相体系是指水相(发酵液或缓冲液)和与水不溶的有机相(单一有机溶剂或混合有机溶剂)构成的体系。水相中含有活性的酶或细胞,有机相中含有甾体化合物。在生物转化过程中,形成油包水和水包油两种情况,底物在水相被酶或细胞催化,所得产物再返回到有机相。由于底物的溶解度在有机相中要远远高于水中,因此可以大大提高投料浓度。由于底物和产物都主要处于有机相中,因此大大降低了底物和产物对酶或细胞的抑制作用。同时,在双液相生物转化体系中,有机相主要作为底物的贮存器和产物的原位分离器,随着生物转化的进行,生成的产物不断被转移到有机相中,水相中的反应平衡向产物方向推动,整个反应的转化率得到提高。选择合适的有机相是双液相系统技术的关键,一方面有机溶剂对生物催化酶或细胞毒性较小,另一方面有机相对底物和产物有较高的溶解度。柏挺[62]在小试发酵规模上,优化大豆油/水双液相系统中分枝杆菌降解植物甾醇为雄烯二酮(AD)的工艺。研究了大豆油在发酵液中的最佳比例,植物甾醇的投料量以及添加表面活性剂对提高植物甾醇转化率的影响,得到最佳转化工艺条件。杨旭锦等[63]提供一种双液相发酵系统发酵分枝杆菌Mycobacteriumnsp.DE6823制备雄烯二酮的方法,在发酵转化完成后,通过膜分离方法,得到含有雄烯二酮的溶液,经浓缩、脱色、过滤、浓缩、重结晶和真空干燥等操作,可得到纯度在99%以上的雄烯二酮。根据选用油相的不同,植物甾醇的转化率为59%~85%。
3.3 双水相体系
双水相体系由两种高聚溶液或者一种高聚物溶液和一种盐溶液构成,并在特定的溶液浓度下形成两种不相容相。微生物细胞被限定在其中一相中,而底物和产物被限定在另一相中。聚合物的存在大大提高了底物和产物在同一相中的溶解度。在反应过程中,底物相中的底物(甾体化合物)会不断地进入微生物细胞所在的相中,经过反应生成产物后再转移回底物相中,这样的双水相体系将底物的浓度提高到接近最大反应浓度,又将产物的浓度降低到很低,减少对转化反应的抑制作用。在双水相体系中,成相介质的选取是非常关键的,它直接决定了底物和产物在两相介质中的分配情况。王艳婷等[64]将分枝杆菌(Mycobacteriumsp.)降解植物甾醇制备雄甾-4-烯-3,17-二酮的研究引入双水相体系中,研究表明,双水相体系具有良好的生物相容性,可用于分枝杆菌对植物甾醇降解、萃取发酵生产雄甾-4-烯-3,17-二酮,可有效缓解转化过程中的产物抑制作用,其工业应用前景良好。
3.4 离子液体体系
离子液体指完全由离子组成、在室温或室温附近温度下成液体状态的盐类,也称为室温熔融盐或室温离子液体。组成离子液体的阳离子主要有烷基咪唑离子、烷基吡啶离子、季铵盐离子等;阴离子主要包括氯化盐类或[BF4]-、[PF6]-和[NTf2]-等。许青青等[65]筛选以离子液体[bmim]Cl为代表,构建了液-液两相体系,结合高效液相色谱分析测定离子液体复合萃取剂对植物甾醇的分配系数和分离选择性。用[bmim]Cl-乙腈/正己烷两相体系萃取,随离子液体浓度的增加,分配系数和萃取选择性系数的增加十分显著,当用20%[bmim]Cl-乙腈萃取时,豆甾醇的分配系数可达7.89,豆甾醇对β-谷甾醇的分离选择性可达65.8,是纯乙腈为萃取剂时的54倍以上。
在生物催化反应过程中,与传统的有机溶剂相比,离子液体具有以下优点:一是为催化体系提供了不同于传统分子溶剂的反应环境,改善生物酶或细胞的活性、提高稳定性,提高反应的选择性和转化率;二是离子液体种类繁多,可选择余地大,酶或细胞溶于离子液体中,反应体系可以循环利用,充分表现了酶或细胞的均相催化效率高、多相催化易分离的优点;三是离子液体难挥发,操作温度范围宽,并且离子液体与有机溶剂不互溶,使得产物易于分离提取。
3.5 超临界流体体系
在超临界流体体系中,物质具备优异的扩散性能和传质效果,酶催化反应发生在接近临界区域内,温度和压力的微小变化就可使溶剂的性质发生独特的变化,可提高酶催化反应的特异性,目前这一技术在医药工业中引起了越来越多的关注。Wang等[66]发现,超临界流体在甾体药物合成起始原料甾醇的萃取中表现优异。赵亚平等[67]采用植物油加工副产品为原料,根据甾醇与其他组分在超临界流体中溶解度的差异,进行纯化精制高纯度甾醇。萃取剂优选CO2,并在萃取过程中加入增效剂醇或酮,得到的甾醇纯度高,得率高,而且不引入有害物质,无环保问题。
3.6 微乳化体系
微乳液是将两种或以上互不相溶的液体经混合乳化后,形成的一种热力学稳定、透明分散体系。在该体系中,生物细胞或酶以分子水平分散于油包水(W/O)状的微乳液中,为酶催化提供大量的油/ 水界面。同时,对于难溶性的甾体化合物,由于其可溶于有机介质,可有效克服传质阻力。微乳液体系可以提高底物的生物利用度,并且可以将细胞和底物分别保留在水相和油相中,从而将细胞毒性降到最低。Zhang等[68]制备了几种不同亲水亲脂平衡的微乳液,并对其稳定性和生物转化效率进行了评价,研究表明微乳液体系中的转化率是传统的水相生物转化系统的3.1倍。由于使用微乳液体系的底物细胞毒性较小,该研究实现了利用再生细胞的微乳液进行重复循环生物转化,6个周期的总产量是传统方法的4.3倍。这种高效的微乳液体系,具有更好的细胞可持续性和生物反应活性,可以扩展到其他不溶性和有毒底物的生物转化。
4 结语与展望
随着微生物技术的不断发展和新型菌株的应用,甾体激素的生物转化不应只强调提高某一步转化效率和专一性,而且要更多地综合应用多种生物技术(例如合成生物学)来提高甾体激素药物生物合成的途径和方法。目前越来越多的具有高转化活性的菌株已经可以通过分子生物学手段对菌株进行改造获得,通过上述甾体药物转化现状的概述,表明转化的成功不仅需要优良菌种,转化体系也至关重要。而为了克服有机溶剂对于环境污染及毒性,不同新型“绿色溶剂”,如离子液体或低溶点共熔溶剂也逐渐应用于甾体药物生物催化体系中,对相关转化方法和转化体系的深入研究,甾体生物转化工业也必然产生新的飞跃。
猜你喜欢
海洋通报(2020年2期)2020-09-04
空间科学学报(2020年4期)2020-04-22
天然产物研究与开发(2018年7期)2018-08-21
天然产物研究与开发(2018年5期)2018-06-13
中成药(2018年4期)2018-04-26
天然产物研究与开发(2018年2期)2018-04-04
中国粮油学报(2017年5期)2017-07-19
中成药(2017年4期)2017-05-17
天然产物研究与开发(2016年1期)2016-06-05
会计之友(2016年7期)2016-04-21
