
季节与小生境对喀斯特灌丛土壤丛枝菌根真菌群落的影响
2019-10-10 06:47赵蕾盖爽爽郑生猛胡亚军刘华英何寻阳
农业现代化研究 2019年5期
赵蕾,盖爽爽,郑生猛,胡亚军,刘华英,何寻阳*
(1. 广西师范大学生命科学学院,广西 桂林 541000;2. 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室,湖南 长沙 410125;3. 中国科学院环江喀斯特生态系统观测研究站,广西 环江 547100;4. 广西科技师范学院食品与生化工程学院,广西 柳州 545000)
喀斯特是一种具有特殊物质、能量、结构和功能的生态系统,由具有溶蚀力的水对可溶性岩石进行溶蚀等作用而形成。喀斯特地区碳酸盐岩充分发育,土壤呈中性至微碱性,土体不连续,土层浅薄[1]。生境泛指生物个体或群落所在的具体地段环境,小生境是在外部形态上可以明显区分、在内涵上具有不同生态有效性的不同小单元[2]。小生境根据微地貌形态共划分为6种类型:石面、土面、石沟、石缝、石坑、石洞[3],喀斯特生境常以石面、土面、石沟、或几种类型组合镶嵌的形式存在。其中石土面及土面以30%岩石裸露率为区分。石土面岩石裸露率大于30%,是类似土面的小型台地[4];土面岩石裸露率低于30%,长宽面积相对较大且土层覆盖均匀,较适宜植物生长,其上分布的植物种类和数量最多,是喀斯特区植被恢复和人工造林的主要利用类型[5]。多样化的小生境也造成土壤养分水分及植物多样化的时空分布格局[6]。西南喀斯特地区存在明显旱雨季,夏季雨量充沛,温度高,湿度大,是植物生长的旺盛季节,植物的生产力高。冬季存在季节性干旱,雨量稀少,温度、湿度低,是大多植物的休眠季节。灌丛在喀斯特地区分布范围广泛,是喀斯特地区石漠化过程中重要的过渡类型,在整个植被演替变化中处于不稳定阶段。生境良好且人为干扰较少的情况下,喀斯特灌丛可能正向演替为森林植被,反之加速岩石裸露的石漠化进程[7]。结合地下微生物调控喀斯特灌丛群落正向演替可能是改善喀斯特脆弱生境的有效途径之一。
丛枝菌根真菌(Arbuscular mycorrhizal fungi,AMF)是生态系统中重要的功能微生物,能与80%左右的陆地植物形成共生关系[8],大量研究表明,接种AMF能提高宿主植物养分吸收,改善水分代谢[9],近年来被开发为脆弱生态环境恢复的生物治理手段[10]。AMF多样性与土壤、气候、及地上植物等环境因子的关系近年来被研究者广泛讨论。Lugo和Cabello[11]研究了3种禾本科的C3代谢植物、2种禾本科及1种蔷薇科的C4代谢植物的宿主物种及其代谢类型、季节变化和放牧对AMF多样性的影响,结果表明AMF的Shannon多样性、丰富度和均匀度没有变化,但菌根真菌侵染密度受季节和宿主代谢途径的强烈影响。Guadarrama等[12]对墨西哥季节性干旱森林干/湿两季土壤AMF孢子进行分离和鉴定以研究AMF丰度、丰富度和多样性的季节变化,结果表明AMF丰富度在干湿地块和季节中存在显著差异,季节变化对AMF多样性有很大影响,但对AMF感染性没有影响。这些相关研究表明季节是影响AMF群落变化的重要因素。但也有研究表明,不论是根际中还是土壤中所含的AMF群落,其组成结构等均不受季节变换的影响[13]。已有研究表明,植物与AMF共生模式在提高喀斯特生态系统稳定性和石漠化治理实践中表现了潜在利用价值[14],但仍缺乏针对喀斯特峰丛洼地灌丛生态系统、不同季节与不同小生境土壤AMF群落研究,而揭示作为喀斯特植被正向演替过渡阶段——灌丛生态系统不同小生境下,土壤AMF的季节变化特征及其影响因子,是合理利用AMF促进喀斯特退化区生态恢复的前提。
1 材料与方法
1.1 研究区概况与样地遴选
研究区位于桂西北典型喀斯特峰丛洼地的广西壮族自治区环江毛南族自治县 (107°51′-108°43′E,24°44′-25°33′N)。属南亚热带向中亚热带过渡的季风气候区。其中南部边缘地带属南亚热带季风气候,中、南部及大、小环江河谷地带属中亚热带谷地气候。全县气候温和,雨水充沛、日照充足、冬暖夏凉、雨热同季、无霜期长。年均气温南部丘陵一带19.9 ℃,北部山区15.7 ℃。全县一月平均气温10.1 ℃,七月平均气温28 ℃;历年最低气温-5.2 ℃,无霜期290 d。全年可照时数4 422 h,全年太阳辐射量98.89 kJ/cm2。年平均降雨量1750 mm,降雨多集中于4至9月份,占全年降雨量的70%;历年最小降雨量922.8 mm,蒸发量1 571.1 mm,空气平均相对湿度79%。经充分勘探后选择了中国科学院环江喀斯特生态系统观测站一处受人为等因素干扰退化,经封育后自然恢复良好的典型喀斯特峰丛坡面灌丛群落,选取植被类型基本一致的1 hm2(100 m ×100 m)面积作为研究样地。
1.2 植被调查与土壤样品采集
在样区内采用Topcon全站仪把样地划分为100个10 m × 10 m相邻的网格样方,经充分勘察后,该样地主要以石土面、土面、石面小生境为主,按传统划分喀斯特异质性小生境的方法[15],基于土被的分布和微地貌特征,划分小生境样区并测量其分布面积,分别于2014年7月(夏季)、2016年1月(冬季)开展土壤样品采样工作,每个样区采用多点(10点)混合取样法组成一个混合样品,采集表层0-15 cm土壤样品约500 g(若土层厚度不到15 cm,则采集至可达深度);采用四分法取土约30 g作为分子试验用样,经液氮速冻后,放置于-80 ℃冰箱保存;植被调查于2014年7月至8月进行。其余土壤样品经自然风干后,用于常规理化指标的测定。最终采集石土面、土面样品数分别为19个和5个,参照5个土面样方面积大小(25-250 m2)对应筛选面积接近的5个石土面样品做后续分析。
1.3 土壤理化性质测定方法
采用pH计测定土壤pH值(水土比=2.5∶1);重铬酸钾容量法-外加热法测定土壤有机碳(SOC);0.5 mol/L的Na2CO3浸提-钼锑抗比色法测定土壤有效磷(AP);中性醋酸铵浸提-火焰光度法测定有效钾(AK);碱解-扩散法测定碱解氮(AN),具体实验操作参照鲍士旦《土壤农化分析》[16]。
1.4 DNA提取、扩增及高通量测序
采用土壤DNA提取试剂盒(Fast DNA®SPIN Kit for Soil,MP)提取微生物DNA。具体的操作步骤严格按照试剂盒说明书执行。提取完成后,分别采用浓度1%的琼脂糖凝胶电泳及核酸检测仪观测扩增样品的片段长度及DNA浓度,并记录每个样品的OD260/280、OD260/230及其DNA浓度含量。确定样品质量完全符合后续试验条件的要求后分装备用。
AMF的18s rRNA扩增采用巢式PCR技术,使用的第1对扩增引物为AML1(5'-ATCAACTTTCGAT GGTAGGATAGA-3')/AML2(3'-GAACCCAAACAC TTTGGTTTCC-5'),目的片段长度在500 bp左右[17];第2对扩增引物是AMV4.5NF(5'-AAGCTCGTAGT TGAATTTCG-3')/AMDGR(3'-CCCAACTATCCCT ATTATCAT-5')引物,目的基因片段长度约 250 bp[18]。以上引物均由上海英潍捷基贸易有限公司合成提供。第1步扩增体系20 μL,包括1 μL DNA模板,上游和下游引物各0.5 μL,2×PCR Ex Taq (TaKaRa, Japan)10 μL,ddH2O 补至 20 μL。程序设定 94 ℃ /5 min,94 ℃ /30 s、58 ℃ /45 s、72 ℃ /1 min, 35 个循环,最后72 ℃延伸10 min。将结束后的PCR产物稀释50倍取1 μL作为第2步DNA模板,扩增体系同样为20 μL,上下游引物各 0.5 μL,2×PCR Ex Taq(TaKaRa,Japan)10 μL,ddH2O 补 至 20 μL。 程 序 设 定 94℃ /3 min,94 ℃ /45 s、60 ℃ /45 s、72 ℃ /1 min,30个循环,最后72 ℃延伸10 min。
高通量测序采用Hiseq2500平台完成,基于Illumina成熟的边合成边测序(SBS)原理进行[19]。HiSeq2500测序与其他高通量测序一样都有其固定的测序引物,在上述第2步扩增体系中,上下游引物5'端添加7个碱基的特定Barcode序列,再对其进行PCR扩增,目的是识别同一批测序数据中不同的样品数据,保证多个样品能够同时上机测序。在将得到的PCR产物按要求保存,送至北京诺禾致源生物信息科技有限公司进行测序。
1.5 数据统计分析
Hiseq高通量数据采用QIIME 2.0和Mothur平台分析。高通量数据质量控制标准:去除碱基质量值(Q值)低于30的序列;去除序列长度小于200 bp,大于250 bp的序列;去除嵌合体及非AMF序列。余下序列以97%相似度进行聚类,提取代表序列在NCBI数据库和MAARJ AM数据库进行物种比对。使用R(vegen包)计算AMF和植被群落多样性指数及MRPP组间差异分析。使用PASW Statistics 18中重复测量方差分析(Within-subjects ANOVA)检验季节和生境对AMF群落丰富度的影响,采用独立配对T检验及pearson相关性分析,分别进行分类间显著差异性及相关性分析。使用Canoco 5.0对AMF群落进行非度量多维尺度分析(Non-metric Multidimensional scaling,NMDS)及其与环境因子的冗余分析(Redundancy ananlysis,RDA)。采用Origin 8.5软件进行后续处理及绘图。
2 结果与分析
2.1 小生境与季节对土壤理化性质的影响
土壤部分理化性质指标在季节与生境之间存在差异(表1)。冬季两种小生境间AK(P=0.000)存在极显著差异;夏季两种小生境间pH(P=0.046)、SOC(P=0.024)存在显著差异;石土面AK(P=0.009)在两种季节间差异极显著;土面AP(P=0.025)在两种季节间差异显著,AK(P=0.003)差异极显著。
2.2 土壤AMF群落组成结构
喀斯特地区灌丛群落含有丰富的丛枝菌根真菌,所有土壤样品测序共得到2 008 658条有效序列,序列平均长度217 bp。AMF序列数1 300 558条,各样品菌根真菌序列数不等(20 315-34 083条),平均约27 095条。筛选最终所得AMF序列总数975 120,分类为154个OTUs,每个样品的OTU个数在93-121之间。数据库中比对后所得AM真菌7科8属,分别为球囊霉科球囊霉属(GlomeraceaeGlomus)、近明球囊霉科近明球囊霉属(ClaroideoglomeraceaeClaroideoglomus)、类球囊霉科类球囊霉属(ParaglomeraceaeParaglomus)、多样孢囊霉科多样孢囊霉属(DiversisporaceaeDiversispora)及雷德克囊霉属(Redeckera)、无梗囊霉科无梗囊霉属(AcaulosporaceaeAcaulospora)、巨孢囊霉科盾巨孢囊霉属(GigasporaceaeScutellospora)、双型囊霉科双型囊霉属(AmbisporaceaeAmbispora)。其中,球囊霉属的相对丰度在两种季节和生境间均居最高,其冬季相对丰度接近90%,夏季约占75%左右。与球囊霉属的季节分布高低趋势相反,相对丰度次之的近明球囊霉属冬季占比低于夏季,类球囊霉属、多样孢囊霉属、无梗囊霉属也呈现同样趋势。双型囊霉属只存在于冬季石土面,其相对丰度仅0.03%。显著差异性分析表明,球囊霉属和多样孢囊霉属在季节间存在显著差异,在生境间仅有盾巨孢囊霉属存在显著差异(图1)。

表1 土壤理化性质Table 1 Physi-chemical properties of studied soils
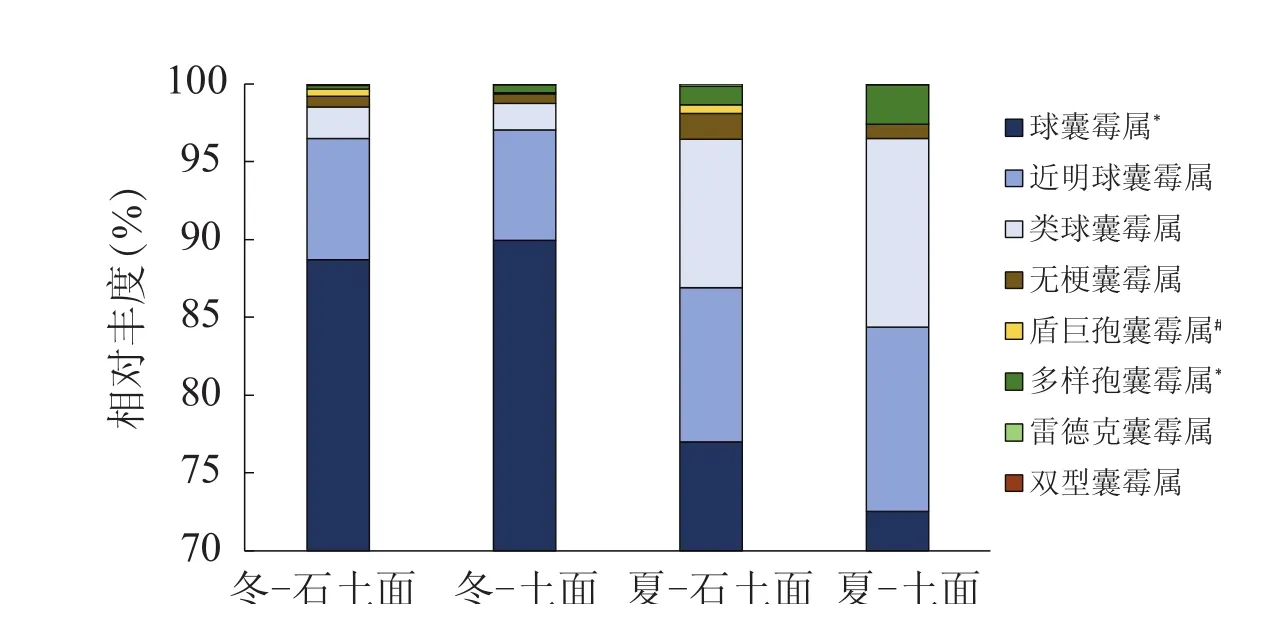
图1 土壤AMF群落属分类水平下相对丰度比较Fig. 1 Comparisons on relative abundance of soil AMF genera between different treatments

图2 土壤AMF群落种分类水平下相对丰度比较Fig. 2 Comparisons on relative abundance of soil AMF taxonomic species between different treatments
为在种的水平反映不同季节及小生境样品间AMF群落物种结构的异同,挑选相对丰度大于1%的OTUs作进一步分析(图2)。结果显示,冬季石土面、土面,夏季石土面、土面各有25、20、23、27个优势种,且优势种相对丰度之和均超80%,其中,有7个优势种相对丰度具有显著的季节变化:OTU25(Glomus Kohout14 A-4;冬季17.77%、夏季9.40%)、OTU3(Glomus sp. VTX00393;冬季5.50%、夏季3.65%)、OTU250(Glomus sp. VTX00202;冬季 3.95%、夏季 2.14%)、OTU53(Glomus Chen14 DL-Glo12;冬 季 2.06%、 夏 季 1.23%)、OTU27(Glomus sp. VTX00120;冬季1.78%、夏季1.21%)、OTU82(Glomus sp. VTX00120;冬 季 1.51%、 夏季0.92%)和OTU32(Glomus sp. VTX00113;冬季1.20%、夏季0.39%)。生境间仅有OTU22(Glomus sp. VTX00064;石土面9.31%、土面3.94%)和OTU119(Glomus GlAd1.2 VTX00288;石土面0.46%、土面0.95%)的相对丰度具有显著性差异。另外,冬季石土面和土面相对丰度最高的种同为OTU25,夏季石土面的则为OTU22,均属于球囊霉属,而夏季土面为OTU6(Paraglomus sp. VTX00001),为类球囊霉属。
测序所得154条OTU中有117条为共有OTUs(图3)。近明球囊霉属OTU108(Claroideoglomus sp. VTX00276)与4种球囊霉属OTU50(Glomus sp. VTX00077)、OTU201(Glomus sp. VTX00085)、OTU212(Glomus sp. VTX00121) 和 OTU343(Glomus Glo16 VTX00120)只存在于冬季;类球 囊 霉 属 OTU4(Paraglomus Alguacil14b Para2 VTX00348)只存在于夏季。OTU213(Glomus sp.VTX00342)为石土面特有,类球囊霉属OTU15(Paraglomus sp. VTX00375)、近明球囊霉属OTU19(Claroideoglomus sp. VTX00193)为夏季石土面特有;双型囊霉属OTU12(Ambispora sp. VTX00283)、OTU73(Ambispora PF27 VTX00283)及多样孢囊霉属OTU99(Diversispora sp. VTX00356)为冬季石土面特有。

图3 土壤AMF OTUs组成Venn图Fig. 3 Venn diagram of AMF OTUs in different seasons and microhabitats
2.3 土壤AMF物种多样性
冬季石土面、土面及夏季石土面、土面实测OTUs个数分别为147,142,139和131,双因素重复测量方差分析结果表明,季节对土壤AMF群落丰富度影响极显著(P=0.007),冬季AMF资源较夏季更丰富,但生境类型、季节与生境类型的交互作用对AMF群落Shannon多样性与均匀度(E)的影响不显著(图4)。
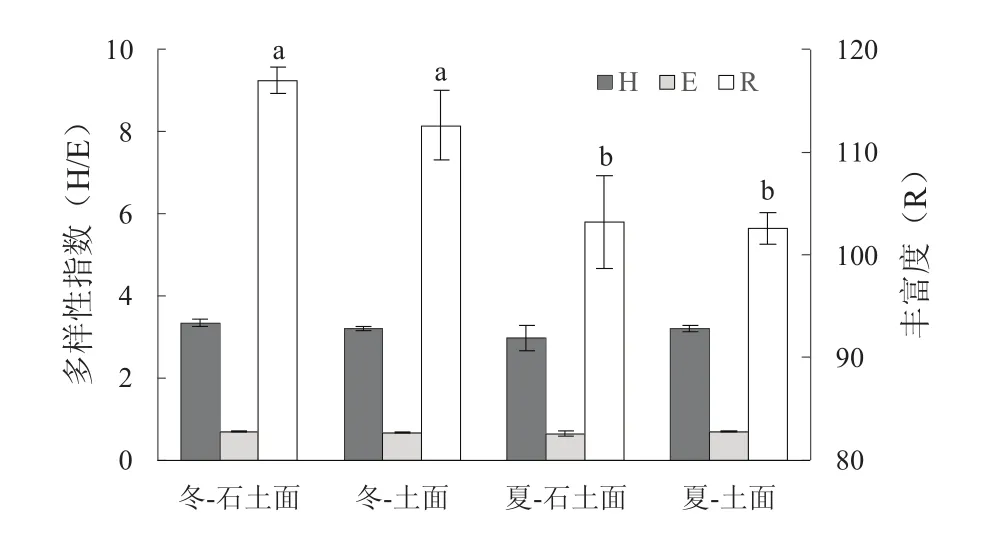
图4 土壤AMF群落多样性Fig. 4 The AMF community diversities in different seasonsand microhabitats

图5 AMF群落特征(A)及其影响因子(B)Fig. 5 The characteristics of AMF community (A) and its Influencing factors (B)
2.4 土壤AMF群落分布的影响因子
NMDS(图5A)结果显示,AMF群落分布存在明显季节差异,冬季群落分布较夏季集中。MRPP分析结果也表明,石土面(P=0.038)、土面(P=0.044)均在季节间表现出显著差异,而在同季节内的小生境间差异不显著。表明季节显著影响AMF群落分布。
以土壤理化指标与植物丰富度为环境因子的RDA(图5B)分析结果显示,第一轴和第二轴的解释量分别为30.76%和15.59%,蒙特卡洛检验依据各指标的解释贡献率排序,表明AP(F=4.0,P=0.012)、AK(F=4.3,P=0.008)及植物丰富度(F=3.8,P=0.002)是影响丛枝菌根真菌群落分布的主要因子(P<0.05),共解释47.30%变异。
3 讨论
3.1 土壤AMF群落对小生境与季节变化的响应
球囊霉属是喀斯特地区丛枝菌根真菌绝对优势种,这与球囊霉属具有较强严峻环境的适应性一致。如内蒙古、宁夏、甘肃盐碱地发现优势种属为球囊霉属[20],在长期受重金属污染的土壤中也广泛分布[21]。另外,球囊霉属在整个研究区的相对丰度占比70%以上,此结果高于多数非喀斯特地区球囊霉属所占比例[22],可能的原因是AMF属于好气性真菌,喀斯特表层土壤有机质含量相对非喀斯特高,且土壤颗粒间隙较大[23],良好的通透性能给AMF带来更好的生存环境。两种季节和小生境下的AMF的Shannon多样性指数和物种丰富度的平均值分别为3.18和108,高于峥嵘等[24]在内蒙古大青山地区的结果,该地区为栗钙土和灰褐土,这可能主要与喀斯特生态系统较强异质性和较高的植物多样性有关。地貌多样性是生物多样性的基础[25],小生境异质性能改善地区生态服务功能,增加物种多样性[26]。由于喀斯特地区地表破碎度高、小生境丰富多样的特殊地质背景,在微地貌水平形成了光热和水分差异的小气候特征,导致土壤理化性质(表1)及植被种类的不同[27]。此外,夏季及冬季的石土面AMF丰富度均较土面的高,推测是因为石土面土层相对土面的薄,水分涵养能力相对较低,且异质性相对更强,从而造成了更多微生态位供土壤AMF共存。
以往研究表明,AMF侵染率、多样性和群落组成均存在季节性变化规律[28-30],也有结果认为AMF群落组成和多样性无季节性差异[31]。本研究中,AMF群落分布呈现显著的季节差异,冬季AMF的Shannon多样性及丰富度(H=3.27, S=114.80)均分别高于夏季的(H=3.09, S=102.90)。AMF依赖宿主植物光合作用提供碳水化合物,不同季节的植物处于不同生长阶段,自身对碳水化合物的需求存在差异,夏季植物为满足地上部分快速生长需要,对碳水化合物和磷的需求增加[32-33],间接影响AMF丰富度和多样性;而冬季植物代谢大大降低,且较低的温度降低植物从土壤中吸收养分的速率[34],同时,喀斯特区冬季季节性的干旱也增加了植物对AMF共生以缓解水分胁迫的需求。
3.2 高异质性喀斯特土壤AMF群落的影响因子
在本研究的各速效养分中,速效磷与速效钾对AMF群落影响极显著。大量研究表明磷是影响AMF群落组成的关键因子[35-36]。魏源等[37]发现土壤磷对贵州茂兰喀斯特森林土壤AMF的定殖和产孢都具有很大影响,不同小生境间这些生态因子,如磷和有机质的显著差异,是导致小生境间AMF多样性指数和物种丰富度差异显著的主要原因。长期以来人们大多关注磷与AMF的密切关系,但近年来,我们的前期研究[38]和其他不少研究表明[39-40],土壤钾素营养对AMF群落及多样性也存在显著影响,因此,土壤钾素营养对于土壤AMF群落组成结构与功能的影响需引起足够的重视。
Martínez-García 等[41]认为,植物物种对 AMF群落组成结构的影响大于土壤养分。在本研究结果中,植物丰富度极显著影响AMF群落结构组成,这与梁月明等[42]的结果一致。植物主要通过其根际分泌物影响根际微生物群落组成结构[43],喀斯特高度空间异质性小生境孕育着不同物种组成的植物群落,而不同物种对土壤AMF具有一定的偏好性。
4 结论
1)球囊霉属(种)是喀斯特灌丛土壤AMF群落优势属(种)。季节是影响喀斯特灌丛生态系统土壤AMF群落的重要环境因素:夏季与冬季土壤AMF群落组成结构差异显著,且丰富度在冬季极显著高于夏季;总体上,小生境类型对土壤AMF群落组成、多样性及丰富度影响均不大,因此利用AMF促进喀斯特峰丛洼地植被恢复可以不考虑小生境的异质性分布。
2)植物丰富度、土壤速效磷和速效钾显著影响喀斯特土壤AMF群落,说明喀斯特土壤AMF存在明显植物群落多样性依赖,且受控土壤有效磷和速效钾供给,因此,营造高植物物种多样性和改善土壤磷钾供给,有望加强AMF促进喀斯特退化植被恢复效应。
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年12期)2021-12-30
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
云南医药(2021年3期)2021-07-21
现代仪器与医疗(2021年2期)2021-07-21
心肺血管病杂志(2020年5期)2021-01-14
科学(2020年3期)2020-11-26
乡村地理(2019年2期)2019-11-16
