
贝莱斯芽胞杆菌(Bacillus velezensis)分类学及功能研究进展
2019-10-09 03:07:38张彩文程坤张欣刘波都海渤姚粟
食品与发酵工业 2019年17期
张彩文,程坤,张欣,刘波,都海渤,姚粟
(中国食品发酵工业研究院有限公司,中国工业微生物菌种保藏管理中心,北京,100015)
贝莱斯芽胞杆菌(Bacillusvelezensis)最早是由RUIZ-GARCA等[1]在西班牙马拉加市的贝莱斯河中发现的。该菌种属于革兰氏阳性菌(G+),菌体呈杆状,大小为0.5 μm×(1.5~3.5)μm,常见单个或成对排列,有时也呈短链排列。芽胞呈椭圆状,位于非肿胀胞囊的中间或末端,不产生伴胞晶体和β-羟基丁酸,可通过鞭毛运动[2],普遍存在于自然界中,如河水[3]、海洋[4]、土壤[5]、植物组织[6]等。目前在NCBI的Taxonomy数据库中显示,贝莱斯芽胞杆菌属于枯草芽胞杆菌群(Bacillussubtilisgroup)分类单元中的解淀粉芽胞杆菌亚群(Bacillusamyloliquefaciensgroup),同时是解淀粉芽胞杆菌植物亚种(Bacillusamyloliquefacienssubsp.plantarum)、甲基营养型芽胞杆菌(Bacillusmethylotrophicus)、“Bacillusoryzicola”的同物异名。在芽胞杆菌属中,“同物异名”现象比较常见,因为早期细菌分类学鉴定技术存在一定的局限,往往缺乏可靠性,特别是枯草芽胞杆菌群和蜡样芽胞杆菌群中的近源菌种不仅在表型上相似,而且在16S rDNA序列上也具有相当高的同源性,因此这些近源菌种之间常出现名称的误用。随着微生物分类鉴定技术的逐渐成熟,一些芽胞杆菌属的菌种也逐步得以正名,其中就包括贝莱斯芽胞杆菌分类学地位的变迁及确定。
芽胞杆菌属(Bacillussp.)包含了许多有特殊功能的菌种,它们在生物农药、生物肥料、生物保鲜、生物降污、食品加工等领域具有特殊的重要性[7]。目前市场上的微生物肥料和饲料中菌种添加剂主要以枯草芽胞杆菌(Bacillussubtilis)为主;苏云金芽胞杆菌(Bacillusthuringiensis)及其伴胞晶体是世界上产量最大的农业微生物杀虫剂,主要防治鳞翅目、双翅目、鞘翅目、螨类等害虫幼虫[8];国家卫生部在2016版《可食用菌种名单》中将能够进行乳酸发酵的凝结芽胞杆菌(Bacilluscoagulans)列入其中。贝莱斯芽胞杆菌是芽胞杆菌属中一种新兴的功能性菌种,已经有很多研究证明,贝莱斯芽胞杆菌在植物促生功能和抗植物病原菌方面能够发挥重要作用。另外,由于其具有将外界一氧化碳物转化为自身营养物质这一特性,甲基营养型芽胞杆菌(即贝莱斯芽胞杆菌)被广泛应用于食品发酵领域,同时在环境保护、工业应用、养殖及医药等领域都具有相应的功能研究[9]。本文对贝莱斯芽胞杆菌的系统分类学地位和功能性研究进展进行概述,以期为进一步研究贝莱斯芽胞杆菌提供参考。
1 贝莱斯芽胞杆菌的分类学地位变迁
1.1 贝莱斯芽胞杆菌的分离和命名
贝莱斯芽胞杆菌最早是从西班牙马拉加市的贝莱斯河河口发现的,RUIZ-GARCA等[1]分别在1999年和2000年利用MY培养基从该河口淡盐水中分离获得CR-14b和CR-502T两株菌,随后通过表型与基因型鉴定手段发现它们是芽胞杆菌属的新种,并于2005年作为一个新的分类单元在IJSEM(International Journal of Systematic and Evolutionary Microbiology)杂志上正式发表,确立了贝莱斯芽胞杆菌的分类地位。在表型上,他们对这2株菌以及其他17株相关芽胞杆菌进行了122项表型测试,选择了92个不同的表型特征进行了聚类分析,发现菌株CR-14b和CR-502T二者之间的表型特征相似性高达94%,而它们与枯草芽胞杆菌枯草亚种(Bacillussubtilissubsp.subtilis)CECT 39T和解淀粉芽胞杆菌(Bacillusamyloliquefaciens)DSM 7T的相似度最高(约90%)。在分子水平上,B.velezensisCR-502T的16S rDNA序列与B.subtilissubsp.subtilisCECT-39T、B.amyloliquefaciensDSM 7T的相似度均大于99%。而B.velezensisCR-502T作为新种的主要依据在于它与相似度高于98%的相关菌种的DNA-DNA杂交(DNA-DNA hybridization,DDH)率均低于20%,其中与B.subtilissubsp.subtilisCECT-39T的杂交率为13.3%,与B.amyloliquefaciensDSM 7T的DNA-DNA杂交率为18.6%。1987年,国际系统细菌学委员会(International Committee on Systematic Bacteriology,ISCB)发表报告指出:通常将DNA-DNA相关性≥70%作为细菌“种”的界限[10],这一标准也被认为是细菌新种鉴定的黄金标准。
1.2 B.velezensis、B.amyloliquefaciens subsp.plantarum、B.methylotrophicus互为同物异名
自B.velezensis命名后的10年里,其分类学地位一直受到人们的关注。在2008年,WANG等[11]选择B.amyloliquefaciensBCRC 11601T、BCRC 14193、BCRC 14711、BCRC 17038与B.velezensisBCRC 17467T进行16S rDNA、gyrB基因序列系统发育分析和DDH相关性分析,根据学名优先权原则,提出B.velezensis是B.amyloliquefaciens(1987年合法化发表)后期的同物异名,主要依据是B.velezensisBCRC 17467T与B.amyloliquefaciensBCRC 11601T、BCRC 14193、BCRC 14711、BCRC 17038的DNA-DNA杂交率分别为74%、89%、74%、76%。在2010年,一株分离于水稻根际土的甲基营养型芽胞杆菌(Bacillusmethylotrophicus)CBMB 205T以新种的名义被发表[12],文章采用16S rDNA系统发育分析确定了8株与CBMB 205T相似性大于98.2%的相关模式菌株,然后将8株模式菌株分别与CBMB 205T进行DNA-DNA杂交,其中与其DNA相关性最高的菌株为B.amyloliquefaciensKACC 12067T(36.0%)。
BORRISS等[13]采用16S rDNA、gyrA、cheA基因序列系统发育分析,结合DNA-DNA杂交技术及比较基因组分析将11株B.amyloliquefaciens分成了两大类,其中基于基因组-基因组距离比较法(genome-to-genome distance comparison,GGDC)计算了模式菌株B.amyloliquefaciensDSM 7T和FZB 42T之间的DNA-DNA相关性大约为77%,同时利用传统的DNA-DNA杂交技术获得二者之间的杂交率为63.7%~71.2%,鉴于DNA-DNA杂交率在70%~80%范围内可用于区分同一种内不同亚种的细菌,他们认为这些结果不足以在种水平区分DSM 7T和FZB 42T,但可以将二者作为B.amyloliquefaciens的2个单独的亚种,并由此提出了以解淀粉芽胞杆菌解淀粉亚种(Bacillusamyloliquefacienssubsp.amyloliquefaciens)DSM 7T(=BCRC 11601T=ATCC 23350T)和解淀粉芽胞杆菌植物亚种(Bacillusamyloliquefacienssubsp.plantarum)FZB 42T(=DSM 23117T)为代表的2个新分类单元。
但在2015年,DUNLAP等[14]提出B.amyloliquefacienssubsp.plantarum应该作为B.methylotrophicus后期的同物异名,因为2个分类单元的模式菌株(FZB 42T和KACC 13105T=CBMB 205T)无论从形态学、生理生化还是核心基因组系统发育分析上都高度相似,并且二者之间基于GGDC方法的DNA-DNA杂交(DNA-DNA hybridization,DDH)率高达85.1%(>70%),核心基因组的平均核苷酸一致性(average nucleotide identity,ANI)值为98.4%(>95%)。对于物种划分而言,一般70%的DNA-DNA杂交率约对应95%的ANI[15]。同年,分离于水稻根部的内生细菌‘Bacillusoryzicola’YC7007与YC7010T以新种的名义在The Plant Pathology Journal杂志上有效发表[16],但该分类名称未在IJSEM杂志上合法化发表。直到2016年,DUNLAP等[17]又进一步对B.velezensis、B.amyloliquefaciens、B.methylotrophicus、B.amyloliquefacienssubsp.plantarum、‘B.oryzicola’的分类学地位重新进行了讨论,他们采用比较基因组学和数字DNA-DNA杂交(digital DNA-DNA hybridization,dDDH)的方法对5株菌B.velezensisNRRL B-41580T(=CR-502T=BCRC 17467T=KCTC 13102T=LMG 22478T)、‘B.oryzicola’KACC 18228T、B.methylotrophicusKACC 13105T、B.amyloliquefacienssubsp.plantarumFZB 42T、B.amyloliquefaciensDSM 7T进行分析,结果显示在核心基因组系统发育树上DSM 7T与其他4株菌位于不同的进化分支,且DSM 7T与NRRL B-41580T、KACC 18228T、KACC 13105T、FZB 42T之间的dDDH值分别为55.5%、55.2%、55.2%、56.2%,而后4株菌两两之间的dDDH值均大于84%,由此指出B.velezensis并不是B.amyloliquefaciens后期的同物异名,他们分别属于2个单独的种,而B.methylotrophicus、B.amyloliquefacienssubsp.plantarum、‘B.oryzicola’应该作为B.velezensis的后期同物异名。
2 贝莱斯芽胞杆菌的分类现状
2017年,FAN等[18]就B.amyloliquefaciens、B.velezensis、B.siamensis提出了一个全新的概念“operational groupB.amyloliquefaciens”:他们首先对芽胞杆菌属(Bacillussp.)中的23株代表性模式菌株的核心基因组进行系统发育分析,从中确定了4个与B.amyloliquefaciensDSM 7T高度相似并具有有效命名的菌种,分别是B.siamensis、B.velezensis、B.methylotrophicus、B.amyloliquefacienssubsp.plantarum;其次在NCBI数据库中获取这4个菌种和B.amyloliquefaciens系列内的66株细菌的全基因组序列进行比较基因组分析,包括rpoB基因系统发育分析、四核苷酸(tetra-nucleotide signatures,TETRA)回归系数、ANI、平均氨基酸一致性(average amino acid identity,AAI)、dDDH、GC mol%、16S rDNA序列相似性分析、核心基因组系统发育分析,其中从rpoB基因以及核心基因组系统发育树上可以清楚地将B.amyloliquefaciens、B.siamensis、B.velezensis/B.methylotrophicus/B.amyloliquefacienssubsp.plantarum分成3组,而且57株B.velezensis/B.methylotrophicus/B.amyloliquefacienssubsp.plantarum系列菌株与DSM 7T的ANI值均在93%左右,而dDDH值均在55%左右,这也进一步证实了之前DUNLAP等[17]的结论;然后他们还分别选择了7株来自于“operational group ‘B.subtilis’”和6株来自于“operational group ‘B.amyloliquefaciens’”的模式菌种,基于它们的ANI和TETRA的中值构建系统树图,结果显示6株“operational group ‘B.amyloliquefaciens’”的模式菌种之间是密切相关的;综上,他们建议将“operational group ‘B.amyloliquefaciens’”作为枯草芽胞杆菌群(Bacillussubtilisgroup)分类等级下的一个新分类单元。另外,部分菌株在分类学命名上存在长期的误用,如芽胞杆菌中的生物防控菌株GB03[19]和QST713[20]经常被当作B.subtilis,但实际上他们是B.velezensis,同时也是“operational groupB.amyloliquefaciens”这一分类单元的成员[18];张欣等[21]采用微生物多相分类鉴定技术对中性蛋白酶生产菌种AS1.398枯草芽胞杆菌(B.subtilis)CICC 10888进行了复核鉴定,发现其分类学地位为解淀粉芽胞杆菌植物亚种(B.amyloliquefacienssubsp.plantarum)。
贝莱斯芽胞杆菌属于枯草芽胞杆菌群,与其近源菌种在表型和16S rDNA序列上难以鉴别,然而随着分子生物学的迅速发展和新技术在微生物分类鉴定中的应用,贝莱斯芽胞杆菌的分类学地位几经波折后终于得以确立。目前“operational groupB.amyloliquefaciens”分类单元中包括B.amyloliquefaciens、B.siamensis、B.velezensis/B.methylotrophicus/B.amyloliquefacienssubsp.plantarum,而根据学名优先权原则,B.methylotrophicus、B.amyloliquefacienssubsp.plantarum、‘B.oryzicola’的当前科学命名应为B.velezensis。
3 贝莱斯芽胞杆菌的功能研究
3.1 农业应用
B.velezensis在农业生产上具有广谱的抗菌活性以及促进植物生长的作用。如表1所示,31株B.velezensis功能性菌株分别分离于河水、农作物土壤、鱼类肠道、植物组织、野牦牛牛粪等不同的基物,其中绝大部分功能研究都与抑制植物病原菌相关,而且B.velezensis常作为农业应用上的生物防控剂和植物生长促生剂。目前,已报道的B.velezensis的抗菌作用机理主要包括:产抗菌蛋白[22]、脂肽类抗生素[23]、由非核糖体多肽合成酶(NRPS)和聚酮化合物(PKS)合成的抗生素[24]、诱导系统抗性[16],同时研究表明B.velezensis具有ACC脱氨酶活性,能够分泌IAA,在促进植物生长的同时,可以提高植物的抗性[25]。如杨胜清等[22]提取了生防细菌B.velezensisS6的发酵液粗提物,经过分离纯化获得抗菌蛋白,并通过菌丝生长速率法和孢子萌发法发现S6的拮抗蛋白对番茄早疫病菌有明显的抑制作用。从受植物病原体侵染的甜菜土壤中分离出的菌株B.velezensisFZB42T,能够产生大量的次级代谢产物,包括bacillibactin、bacillomycin D、bacilysin、difficidin、fengycin、macrolactin、surfactin,其全基因组序列分析表明,有9个NRPS和PKS基因簇直接与生物活性肽和聚酮化合物的合成相关,其中有3个基因簇(mln、bae、dfn,基因长度共199 Kb)直接合成具有抗菌作用的聚酮类化合物:macrolactin、bacillaene、difficidin[26]。分离于水稻根际的B.velezensisCAU B946[27]能够生成脂肽类、聚酮类化合物(PKS)、芽胞杆菌溶素等抗生素,并且在控制烟草黑胫病、水稻纹枯病、棉花枯萎病、棉花黄萎病和小麦疮痂病等植物病害上显示出了巨大的潜力;从小麦根际分离的YAU B9601-Y2[28]同样能够产生bacillomycin D、fengycin和difficidin;菌株B.velezensisM75分离于用于蘑菇栽培的废棉,对植物病原真菌具有明显的拮抗作用[24];来源于野牦牛牛粪的菌株LM2303也能够分泌很多抗菌性次级代谢产物,可以有效抑制禾谷镰刀菌的分生孢子萌发和菌丝生长[29-30]。而且,菌株LM2303、CAU B946、YAU B9601-Y2和M75的全基因组序列分析中均发现了与FZB42T相似的9个具有抗菌功能的NRPS和PKS基因簇,此外,在YAU B9601-Y2中发现了负责合成和修饰mersacidin(一种B型羊毛硫抗生素)的完整基因簇,但在与B.velezensis物种亲缘关系比较近的其他菌株中未观察到[24]。而菌株M75和LM2303含有kijanimicin(氯丝菌素)操纵子片段,使它们能够对一些革兰氏阳性菌和厌氧微生物产生抗性[24,30]。
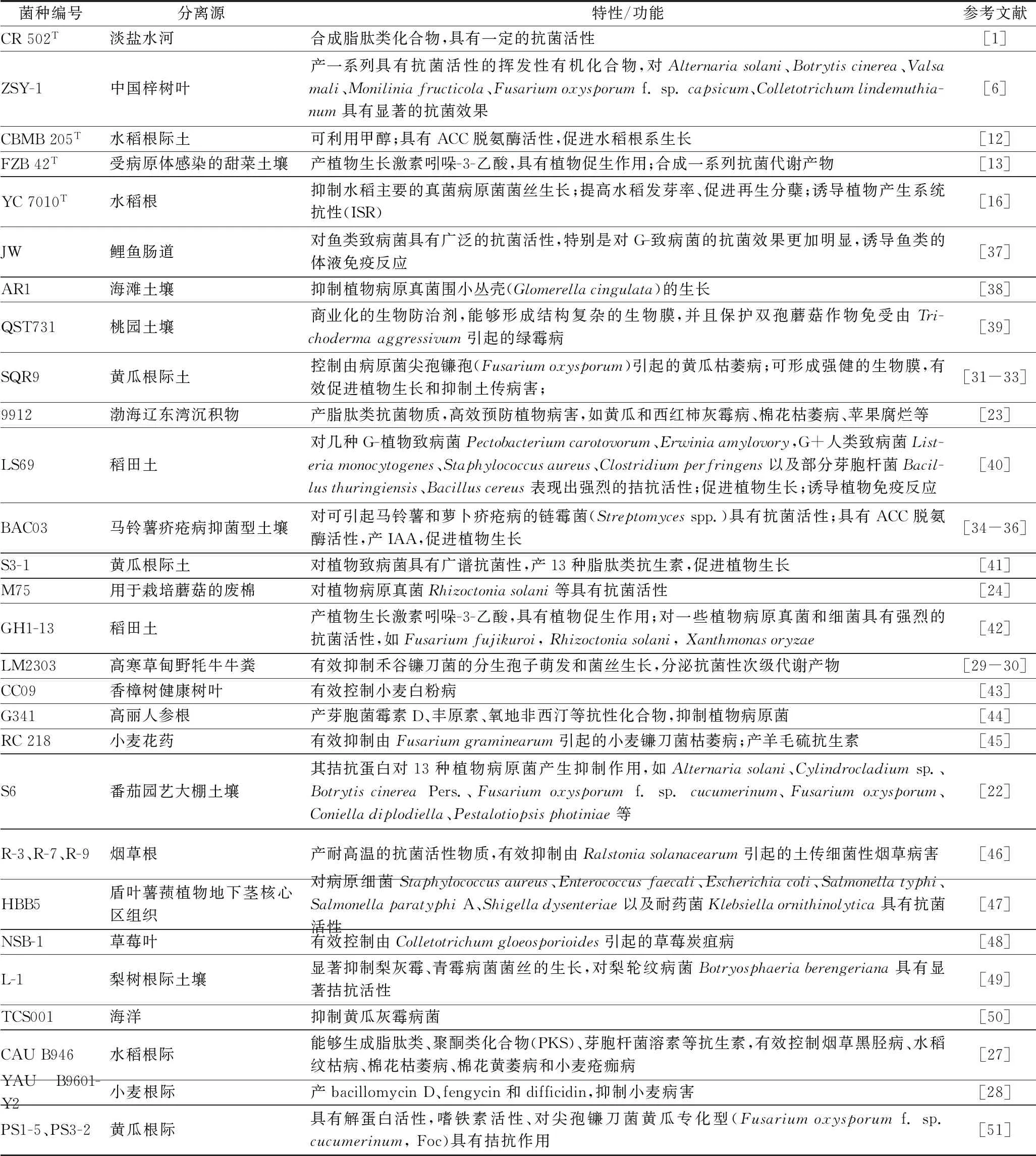
表1 B.velezensis不同菌株分离源及功能特性统计Table 1 Habitats and functions of different B.velezensis strains
由于不同菌株具有不同的基因组特征,因此这些菌株的抗菌活性和次级代谢产物也有可能不同。菌株B.velezensisSQR9(曾用名B.subtilisSQR9[31]、B.amyloliquefaciensSQR9[32])是一株分离于黄瓜根际土的广谱性拮抗菌,常作为农作物生产中的生物肥料和生物防治剂,能够形成强健的生物膜,具有突出的根系定殖能力,有效促进植物生长和抑制土传病害[33]。来源于马铃薯疥疮病抑菌型土壤的B.velezensisBAC03(曾用名B.amyloliquefaciensBAC03[34]),能够有效抑制由Streptomycesscabies引起的马铃薯和萝卜疥疮病[35]。为了确定BAC03促进植物生长的功效,MENG等[36]测试了菌株BAC03对9种类型植物(包括甜菜,胡萝卜,黄瓜,胡椒,马铃薯,萝卜,南瓜,番茄和萝卜)的潜在促生作用,结果表明菌株BAC03具有ACC脱氨酶活性和分泌IAA的能力,对9种植物都具有一定的促生作用。
3.2 工业应用
甲基营养型芽胞杆菌(即贝莱斯芽胞杆菌)C412的胞外分泌物具有絮凝作用,对高岭土悬浊液的絮凝活性高达98.27%,微生物絮凝剂的产率约为4.25 g/L,该微生物絮凝剂的主要组分是多糖和极性官能团,具有较好的耐热性和耐酸碱性,具有潜在的工业应用价值[9]。另外,由于甲基营养型芽胞杆菌具有将外界一氧化碳物转化为自身营养物质这一特性,被广泛应用于食品发酵领域[9]:从自然发酵的豆豉中分离获得的甲基营养型芽胞杆菌B-1具有较高的酶活性,接种于豆粕中进行固态发酵时能够高效分解豆粕中的大分子蛋白质[52];从酱香型大曲中分离获得的甲基营养型芽胞杆菌FBKL 1.0190产中性蛋白酶、糖化酶、纤维素酶和脂肪酶的能力非常突出,对提高酱香型大曲的品质具有良好的应用前景[53]。从土壤中分离的甲基营养型芽胞杆菌SK19.001能够不依赖谷氨酸发酵生产无毒的γ-聚谷氨酸,可直接应用于食品工业等领域[54]。
3.3 环境保护应用
在环境保护方面,贝莱斯芽胞杆菌也发挥着一定的作用,然而绝大数相关研究同样是以甲基营养型芽胞杆菌的命名展开的。最初,RUIZ-GARCA等是为了筛选出产表面活性剂的新菌种而分离到了贝莱斯芽胞杆菌CR-14b和CR-502T,而事实也证明了这2株菌具有作为生物表面活性剂的潜力。目前的生物表面活性剂在污水处理[55]、生物修复、重金属提取以及油泥预处理等方面都有广泛的应用[56]。如从复合重金属污染场地分离的甲基营养型芽胞杆菌JY-04能够高效吸附水体环境中的重金属,而且能够在土壤中强化植物提取重金属的能力[57];谭泽文等[58]从土壤中筛选出的一株甲基营养型芽胞杆菌RF2具有餐厨垃圾防蝇产蛆效果,这可能与该菌在生长繁殖阶段分泌出大量的抗生素类或抗菌肽类物质有关。荣璐阁等[59]通过使用表面活性剂吐温80(Tween80)和鼠李糖脂(RL)强化了甲基营养型芽胞杆菌对柴油土壤的修复效果。
此外,贝莱斯芽胞杆菌还可以作为饲用微生物菌剂,同时有望在医疗保健方面发挥更重要的作用[9]。同一种不同株B.velezensis在功能上存在着差异,因此还需要进一步发掘自然界中B.velezensis的新菌株资源,以开发利用新功能菌株。
4 展望
近年来,由于微生物分类鉴定技术的迅速发展,微生物分类系统的更新速度也不断加快,不仅新种的数量不断增长,一些微生物的分类学地位也发生了变化,贝莱斯芽胞杆菌所属的枯草芽胞杆菌群即是分类变化快速的典型代表[60]。随着分子生物学的迅猛发展,特别是对基因组学研究的兴起,进一步完善了微生物的分类鉴定方法和标准。贝莱斯芽胞杆菌分类学地位的变迁及确立经历了一个相对曲折的过程,其后期的同物异名菌种出现的原因在于:一方面是所采用的细菌分类鉴定方法存在局限性;另一方面是已报道的同物异名现象对后期相关菌种的研究存在一定程度上的制约性。因此未来对于微生物新种的发表,建议在多项分类技术的基础上,与基因组学方法相结合,互为补充和验证,从而让菌种鉴定获得更科学、更准确的结论。
贝莱斯芽胞杆菌在农业、工业、环境保护、养殖等众多领域都有相应的功能研究,其中在农业推广上的运用最多,主要体现在其对多种植物病原微生物的广谱抗性以及促生作用。在过去几个世纪,化肥和农药在提高作物质量和产量方面发挥了非常重要的作用,但同时对地球环境、人类健康及生态系统的稳定性都构成了潜在的威胁,因此使用生物控制剂防治植物病原菌不失为一种更安全且可持续的战略措施[61]。目前,芽胞杆菌Bacillusspp.已经成为农业应用上非常广泛的一类生防细菌,它们能够产生芽胞,具有种群庞大、繁殖力强、抗逆性强、理化性质稳定、产物丰富等特点[62-63]。本文中已报道的贝莱斯芽胞杆菌(B.velezensis)来源于不同的生境,说明其在自然界中广泛存在,而且可以通过产抗菌性次级代谢产物、诱导抗性、分泌IAA等方式对多种植物病原微生物产生广谱抗性,促进植物生长。因此,开发利用贝莱斯芽胞杆菌(B.velezensis)作为生防制剂具有良好的农业应用前景。为了更好地利用贝莱斯芽胞杆菌控制植物病害,需重视特殊生境中的B.velezensis优良菌株的筛选,加强菌剂研发工作;利用分子生物学、基因组学和蛋白组学等方法明确生防相关基因在植物、病原微生物、贝莱斯芽胞杆菌三者之间发挥作用的机制;可考虑改良B.velezensis的功能基因,提高其抗菌活性、诱导抗性,增强抑菌广谱性等,以期尽早实现贝莱斯芽胞杆菌菌剂的商品化、规模化生产。另外,同物异名甲基营养型芽胞杆菌(B.methylotrophicus)在食品加工、工业等方面的应用已经有相对成熟的研究,这为今后进一步开发B.velezensis在其他领域的功能提供了新的思路。
猜你喜欢
流程工业(2022年11期)2022-11-30 06:51:12
中国生物防治学报(2022年3期)2022-07-09 10:00:22
军事文摘(2021年18期)2021-12-02 01:28:04
微生物学杂志(2021年2期)2021-07-01 11:01:06
微生物学杂志(2020年2期)2020-12-31 07:17:13
疯狂英语·新策略(2019年8期)2019-10-18 00:59:28
实用手外科杂志(2015年3期)2015-08-27 01:53:08
山西农经(2015年7期)2015-07-10 15:00:57
食品工业科技(2014年15期)2014-03-11 18:17:52
教育与职业(2014年1期)2014-01-14 02:56:34
