
代谢组学在白血病中的研究进展
2019-09-25 07:39宋志强朱臻宇柴逸峰
药学实践杂志 2019年5期
宋志强,朱臻宇,刘 悦,柴逸峰
(海军军医大学药学院,上海 200433)
白血病是造血干细胞在各个分化阶段由于分化阻滞、凋亡障碍并克隆增生引起的血液系统恶性肿瘤性疾病,根据白血病细胞类型和成熟程度,可将白血病分为4种类型:急性髓细胞性白血病(acute myeloid leukemia,AML)、慢性髓细胞性白血病(chronic myeloid leukemia,CML)、急性淋巴性白血病(acute lymphoid leukemia,ALL)和慢性淋巴性白血病(chronic lymphoid leukemia,CLL)[1]。在中国,白血病发病率为3/10万~4/10万,是儿童中最常见的恶性肿瘤,约占所有恶性肿瘤的30%[2]。致死率在恶性肿瘤中分别排第6位(男性)和第8位(女性),在儿童中则位于首位。
目前,白血病的诊断主要依赖于血液和骨髓检测,以获得白血病的形态学、免疫学和细胞遗传学信息。然而,在白血病早期和缓解期,简单的血液检查并不能发现白血病。所以,白血病的早期诊断主要依靠骨髓穿刺获得细胞形态学和免疫表型等信息[3]。但这一侵入性的检查方式给患者(特别是儿童)带来了极大的身体创伤和心理创伤。同时,由于白血病的隐匿性、非特异性和复杂性[4],许多白血病患者并没有得到早期诊断,以致错过了早期治疗的时机。因此,有必要寻找潜在的生物标志物用于白血病的早期预警和诊断。近年来,随着白血病化疗效果的提升,白血病患者的5年生存率达到了54%,1~4岁儿童ALL患者的生存率更是高达95%[5-6]。但是,停止治疗后,约有20%的患者复发[7],耐药是其中的一个重要原因。因此,一方面,需要探索耐药机制,克服耐药难题,另一方面,应当开发新药并研究药效机制。虽然目前已有许多关于白血病危险因素的研究[8-9],但其发病机制尚不完全清楚,因此需要运用新的方法加以探索。
内源性小分子代谢物可以很好地反映机体的代谢状况,已被广泛用于多种疾病的发病机制、诊断方法和药效机制研究[10-11]。白血病患者由于代谢重编程,其内源性小分子代谢物也会发生变化,因此,可以通过检测内源性小分子代谢物的变化探索白血病的发病机制、诊断方法和药效机制,这无疑为白血病研究提供了一个新的视角。
代谢组学是继基因组学、蛋白质组学和转录组学后,于20世纪90年代中期兴起的一门学科,其通过探究机体在生理或病理情况下产生的差异性内源小分子代谢物研究不同疾病的发病机制和规律,并依此提供疾病治疗的新策略[12]。代谢组学研究的对象是相对分子质量<1 000的内源性代谢物,通过对它们实时动态的定性或定量分析,可以从整体角度更直观地了解机体的功能变化,并与临床症状相联系[13]。总之,代谢组学能够将高通量、高分辨率的分析技术与生物信息学相整合[14],高效、全面、无差别地寻找内源性小分子代谢物。本文主要综述代谢组学在白血病发病机制、诊断方法和药效机制中的研究进展。
1 代谢组学
根据分析目的不同,代谢组学可分为靶向代谢组学[15]、非靶向代谢组学[16]和伪靶向代谢组学[17]。靶向代谢组学用于发现特定的代谢物,非靶向代谢组学用于尽可能多地发现代谢物,伪靶向代谢组学则结合了两者的优点,拥有更好的重复性和更大的线性范围。目前,代谢组学广泛使用的分析方法有质谱(MS)与核磁共振技术(NMR)[18]。同时,MS常与气相色谱(GC)[19]、液相色谱(LC)[20]、毛细管电泳(CE)[21]等技术联用。几种技术之间各有优势和适用范围。MS拥有高灵敏度、高特异度和高选择性的特点,但同时有样品处理过程复杂、受基质效应和离子化程度的影响等不足。NMR能够对样品实现无创、无偏向的检测且样品前处理简单,但灵敏度和特异性低,常需要大样本量分析。而色谱和电泳技术虽然定性能力不足,却具有高通量和高分离度的特点[22]。为了实现代谢组学全面、高效、高通量的优势,常将多种技术进行联用,这也是今后代谢组学发展的重要趋势。通过分析技术获得代谢物数据信息后,需要对数据进行处理,通常使用单变量和多变量的统计分析方法获得差异代谢物。单变量分析方法常用的有参数检验与非参数检验,多变量分析包含主成分分析(PCA)、偏最小二乘法(PLS-DA)与正交偏最小二乘法(OPLS-DA)。其中,PCA又称为非监督学习方法,PLS-DA与OPLS-DA被称为监督学习方法。运用这些分析方法,可以很好地对代谢轮廓进行分析。近年来,本课题组利用代谢组学对疾病进行研究已经取得了一些研究成果。Liu等[23]运用GC/MS与LC/MS探究蓝萼甲素对肝癌细胞的作用机制,发现用蓝萼甲素治疗后,癌细胞的葡萄糖、脯氨酸、乳酸盐等内源性小分子代谢物发生变化,蓝萼甲素可能通过嘌呤、嘧啶、鞘脂和氨基酸代谢发生药效作用。Wu等[24]利用 UHPLC-QTOF/MS对严重尿毒症皮肤瘙痒和轻微尿毒症皮肤瘙痒患者的血清进行了代谢组学研究,发现差异主要是溶血性磷脂酰胆碱、苯乙酸、亚牛磺酸等内源性小分子代谢物,主要涉及磷脂代谢等代谢通路,说明磷脂代谢在尿毒症皮肤瘙痒中具有重要的调节作用。
2 内源性小分子代谢物在白血病发病机制中的研究
Mayer等[25]用蛋白质组学和代谢组学探究衰老导致慢性B淋巴细胞性白血病(B-CLL)的分子机制。通过年轻人、老年人的正常B淋巴细胞与B-CLL细胞的对比发现,衰老可以使细胞的活性氧含量显著增高,而与线粒体氧化应激相关的蛋白含量也明显提高。与正常B淋巴细胞相比,B-CLL的谷氨酰胺消耗增加,脂肪酸的β氧化增强。为了验证以上发现,Mayer等进一步运用LC-MS对4例老年B-CLL患者和5例健康志愿者的B淋巴细胞进行靶向代谢组学分析,发现谷氨酸和谷氨酰胺含量降低,谷氨酰胺的分解代谢物丙氨酸含量增高,这证明B-CLL细胞谷氨酰胺分解增强,与上述研究相一致。同时,在癌细胞中,谷氨酰胺分解与蛋白质、脂肪酸合成及活性氧产生有关[26]。这些与谷氨酰胺相关的内源小分子代谢物变化说明衰老导致的线粒体氧化应激和代谢重编程与B-CLL的发生密切相关。
Agathocleous等[27]运用LC-MS对小鼠骨髓来源的CD150+CD48-LSK造血干细胞(HSCs)、CD150-CD48-LSK多功能祖细胞(MPPs)与多种功能受限的造血祖细胞进行代谢组学分析,发现维生素C含量在功能受限的造血祖细胞中降低,进一步研究发现维生素C水平降低加速了白血病的发生,而维生素C在造血干细胞的累积则可抑制白血病的发生。这说明内源性小分子代谢物维生素C在白血病的发生中具有重要作用。
3 内源性小分子代谢物在白血病早期诊断中的研究
癌细胞的生长需要大量葡萄糖提供能量,而且即使在氧供充足的情况下依然选择糖酵解的方式供能,被称为Warburg效应[28]。一方面,白血病细胞可以通过增强糖酵解促进其他生物大分子的合成[29];另一方面,糖酵解产生的能量远低于糖的有氧氧化,所以白血病细胞需要通过增强糖异生满足对糖的需求。在代谢应激的情况下,白血病细胞发生代谢的重编程,从而导致代谢物种类或含量的改变,因此,用代谢组学分析小分子代谢物的变化有利于白血病的早期诊断。
Musharraf等[30]将气相色谱与三重四极杆质谱技术(GC-QQQ-MS)联用,对72例ALL患者、64例AML患者、50例再生障碍性贫血(APA)患者和76例健康志愿者血清进行了代谢组学分析。运用方差分析(ANOVA)从检测到的1425个代谢物中发现了27种显著差异代谢物(与健康组相比,P≤0.001且变化倍数>3),白血病患者的软脂酸、硬脂酸等代谢物含量下降,花生酸、油酸等代谢物含量升高,并运用PLS-DA 建立了一个基于这27种代谢物的APA、 ALL、AML与健康患者的区分模型,模型的敏感度和特异性分别是91.42% 与90.76%。说明这些脂肪酸代谢中的内源性小分子代谢物可以很好地用于急性白血病的早期诊断。进一步研究发现,ALL组与其他3组的代谢物差异最大,而APA与AML组的差异最小,这从另一个角度说明APA可以发展为AML[31]。
Yang等[32]应用气相色谱与质谱联用技术(GC-MS)对26例新发CML患者,26例经酪氨酸激酶抑制剂(TKI)治疗好转的CML患者和26例健康志愿者的血清进行了代谢组学研究。经过t检验和PLS-DA等方法发现代谢特征并成功发现了6种CML患者与健康志愿者之间的显著差异物,分别是CML血清中降低的肉豆蔻酸、甘油,升高的肌醇、半乳糖、乳酸、甘氨酸(VIP>1,P<0.05,AUC>0.7)。并且这些差异代谢物在经TKI治疗后均有所回调。PLS-DA预测模型的可解释性(R2X=30.3%,R2Y=86.1%)与可预测性(Q2=0.748)说明了该模型的可靠性。它们之间的代谢差异主要发生在半乳糖代谢、丙酮酸代谢、甘油酯代谢、甘氨酸/丝氨酸/苏氨酸代谢中。这6种内源性小分子代谢物可以作为CML早期诊断的生物标志物。
Chen等[33]应用气相色谱-飞行时间质谱(GC-TOF-MS)对400例新发AML患者和446例健康志愿者的血清进行了代谢组学研究,经过Wilcoxon秩和检验和OPLS-DA等模式识别方法研究发现,与健康志愿者相比,AML患者的血清中3-磷酸甘油、乳酸、柠檬酸含量降低,丙酮酸、2-氧化戊二酸和2-羟戊二酸水平升高。Chen等进一步用这6个代谢标志物构建了AML的预后风险评估(PRS)体系,PRS评分低的白血病患者生存率低。通过比较PRS评分低与PRS评分高的白血病患者AML原始细胞的基因表达模式,发现在PRS评分低的患者中糖酵解与三羧酸循环的基因表达水平显著增加。同时,体外实验结果表明糖酵解增强会降低白血病细胞对抗白血病药物阿糖胞苷的敏感性,而抑制糖酵解则可以抑制AML的细胞增殖并增强阿糖胞苷的作用。综上,利用这6种显著差异代谢物可以对AML进行早期诊断和预后分析,还可以考虑将糖酵解抑制剂与化疗药合用治疗AML。
Wang等[34]应用氢谱核磁共振(1H-NMR)对183例新发AML患者和232例健康志愿者的血清进行了代谢组学研究。将AML患者按照外周血或骨髓白血病细胞的核型分为58例低危、119例中危和6例高危AML患者,并与健康志愿者的血清进行对比。经过ANOVA与OPLS-DA等模式识别方法发现代谢特征并成功建立了不仅能够区分健康志愿者与患者,还能区分低危与中危AML患者的模型。AML患者血清中苯丙氨酸、葡萄糖等小分子代谢物含量升高,胆碱、高密度脂蛋白等含量下降。与低危组患者比较,中危组患者血清中异亮氨酸、谷氨酸、胆碱和高密度脂肪酸等代谢物含量上升,而低密度脂肪酸和极低密度脂肪酸含量下降。基于1H-NMR 的代谢组学方法检测到的小分子代谢物可以用于白血病的早期诊断和病情严重程度的判断。
综上,由于白血病细胞的代谢重编程,可以利用代谢组学发现白血病的内源性小分子代谢物,从而实现白血病的早期诊断,现将以上研究总结,见表1。
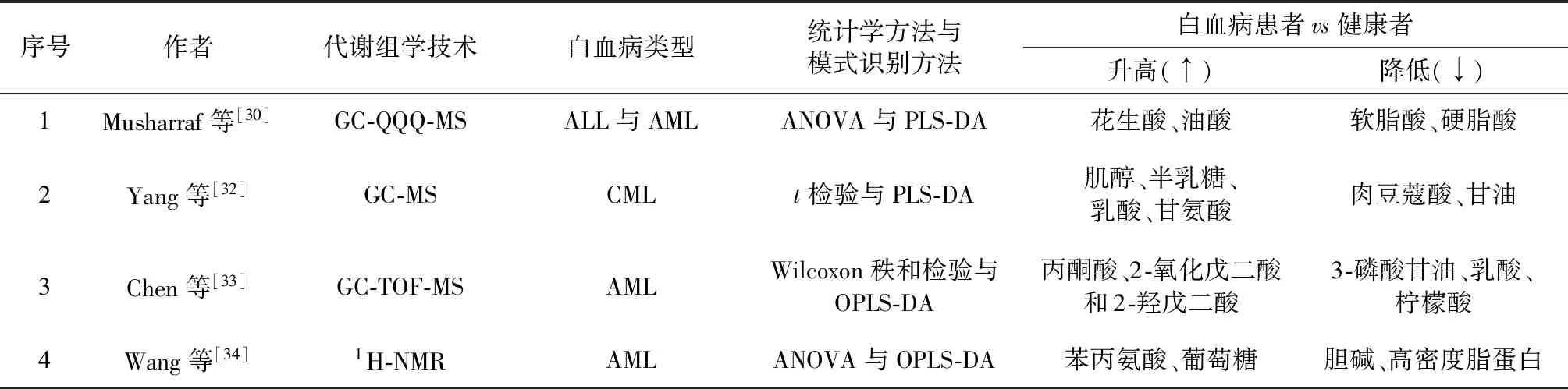
表1 内源性小分子差异代谢物在白血病早期诊断中的应用举例
4 内源性小分子代谢物在白血病药效机制中的研究
Zhang等[35]运用HPLC-MS进行雷公藤红素治疗急性早幼粒白血病(APL)的机制研究,通过对照组与不同浓度雷公藤红素干预的APL 细胞HL-60的细胞代谢组学分析发现,尿苷是对照组与治疗组的显著代谢差异物,且随着雷公藤红素浓度的增高,尿苷含量呈下降趋势。进一步研究发现,尿苷可以抑制细胞凋亡,尿苷的缺乏可以促进雷公藤红素诱导的线粒体凋亡。因此,雷公藤红素可以通过降低尿苷含量发挥促进HL-60凋亡的作用。
二甲双胍常用作治疗2型糖尿病,近年来研究表明,二甲双胍可以通过抑制线粒体复合体Ⅰ使得一磷酸腺苷(AMP)水平升高,进一步影响能量代谢,从而抑制肿瘤细胞的生长[36-37]。Scotland等[38]用液相色谱-线性离子阱/静电场轨道阱组合式高分辨质谱(LC-LTQ Orbitrap MS)对AML细胞内代谢物进行了代谢组学研究。发现使用二甲双胍后,糖酵解的上游代谢产物(6-磷酸葡萄糖、6-磷酸果糖、1,6-二磷酸果糖)、磷酸戊糖途径的氧化产物和肉碱含量下降,乙酰肉碱和嘌呤增加。进一步研究发现,二甲双胍在抑制线粒体复合体Ⅰ电子转运的同时,能够通过增强糖酵解与脂肪酸代谢抑制白血病细胞的增殖。这些内源性小分子代谢物为白血病药效机制研究提供了一个新的视角。
苯扎贝特(BEZ)在临床上常用于治疗高血脂症[39],Southam等[40]利用13C标记的葡萄糖与直接进样傅立叶变换离子回旋共振(FT-ICR)质谱技术来测量AML细胞系和伯基特淋巴瘤细胞系中脂质组代谢与脂质合成的变化,从而研究BEZ 和醋酸甲羟孕酮(MPA)合用治疗白血病和淋巴瘤的机制,研究发现其可以通过减少硬脂酰辅酶A去饱和酶1(SCD1)含量和单链不饱和脂肪酸的合成发挥抗癌作用。这是临床上首次报道靶点是SCD1的治疗白血病和淋巴瘤的药物。
Stäubert等[41]利用RNA测序、非靶向代谢组学方法和同位素标记质谱分析技术对过量表达P-糖蛋白的急性T淋巴细胞白血病耐柔红霉素机制进行了细胞代谢组学研究,发现耐药白血病细胞泛酸的吸收能力下降,对糖分的需求增加,更多地依靠糖酵解和三羧酸循环供能,脂肪酸氧化和谷氨酰胺分解降低。这项研究促进了急性T淋巴细胞白血病耐药的机制研究。
5 讨论
目前,代谢组学主要通过色谱、质谱与核磁共振技术以及它们之间的联合使用检测出白血病的代谢物,然后经后续的模式识别方法和统计分析寻找出显著性内源小分子代谢物,进一步找出相关代谢通路,从而实现对白血病的发病机制、诊断和药效机制的研究。在白血病发病机制研究方面,通过代谢组学技术与其他分子生物学技术合用,揭示了氧化应激和维生素C在白血病发病机制中的重要作用。从现有的研究可以看出,白血病患者的代谢重编程主要涉及脂肪酸代谢、重要氨基酸代谢和糖酵解与糖异生过程,其中的内源性小分子代谢物可以辅助白血病的早期诊断和病情判断。另外,内源性小分子代谢物还可以用于探索雷公藤红素、二甲双胍与BEZ对白血病的药效机制研究,白血病对柔红霉素的耐药机制也逐渐清晰。由于代谢组学分析的样本通常是血液和细胞,因此避免了因骨穿这一侵入性检查方式造成的心理和身体创伤,以一种微创的方式实现白血病的早期诊断和机制研究是代谢组学的一大优势。同时,相对于基因组学和蛋白质组学,代谢组学检测的是基因、蛋白等综合因素作用下的最终代谢物变化,比其他组学更能综合、全面地反映白血病患者的生物状态。代谢物种类也远少于基因与蛋白的数目,研究相对简单。
随着代谢组学的发展,代谢组学越来越多地与基因组学、蛋白质组学等相结合,利用各组学之间的优势共同探讨白血病的发生发展。同时,代谢组学技术也常与RNA测序、同位素标记等分子生物学技术一起使用,利于对白血病进行深层次的研究,找出更有意义的显著的差异代谢物,促进白血病发病机制、早期诊断、药效机制等方面研究的发展。这种多组学、多技术合用的研究方式,也是代谢组学未来发展的一个重要趋势。
虽然利用代谢组学在白血病研究方面取得了一些成就,但仍存在一些待改进之处:①由于代谢组学起步较晚,代谢组学数据库仍需要进一步完善,代谢组学的数据处理和分析方法需要进一步发展,这样更有利于准确分析代谢通路,探究白血病发生机制。②目前虽然已经发现了白血病中的多种差异代谢物,但是不同的文献中发现的潜在标志物及涉及的代谢通路差异较大,仍需整合多方面的资源信息构建全面的白血病代谢网络。③目前的研究样本大多以血液或细胞为分析对象,如果能够将脑脊液与尿液等更多的生物样本一同综合分析,将能够更好的表征由白血病引起的代谢变化。④虽然应用代谢组学发现了多种与白血病密切相关的内源性小分子代谢物,但缺乏临床大样本研究验证其可靠性,与实际运用还有一定的差距。
猜你喜欢
现代临床医学(2022年4期)2022-09-29
昆明医科大学学报(2021年8期)2021-08-13
科学(2020年3期)2020-11-26
中国医院用药评价与分析(2020年3期)2020-05-29
中国循证心血管医学杂志(2020年11期)2020-01-08
中成药(2019年12期)2020-01-04
农药科学与管理(2019年5期)2019-08-13
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医学装备(2016年6期)2016-12-01
现代养生·下半月(2015年6期)2015-09-07
